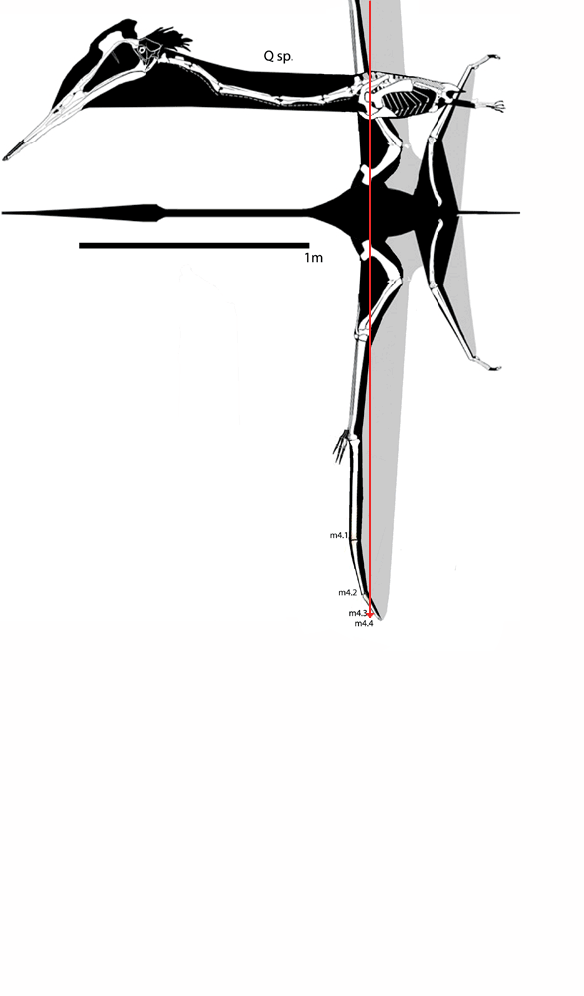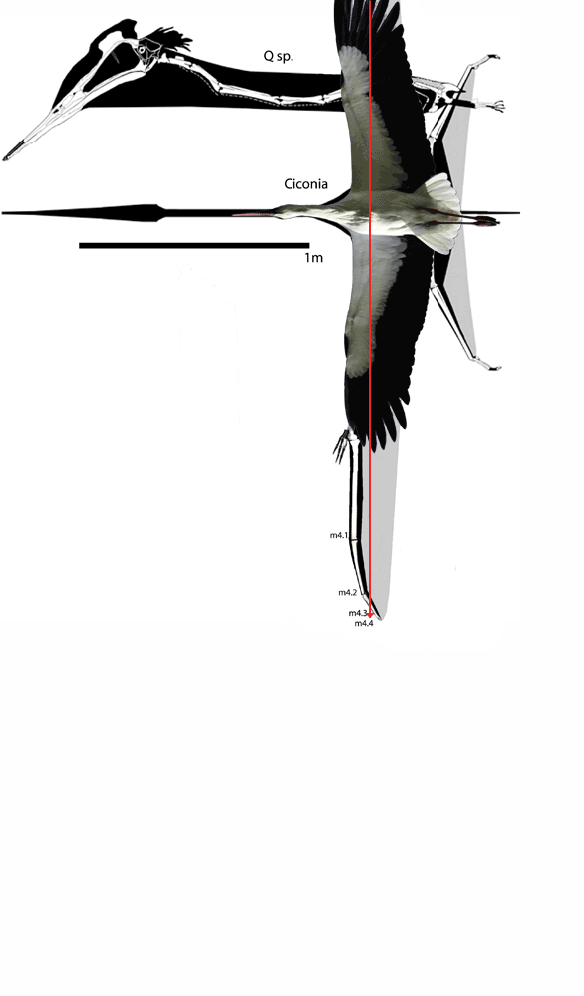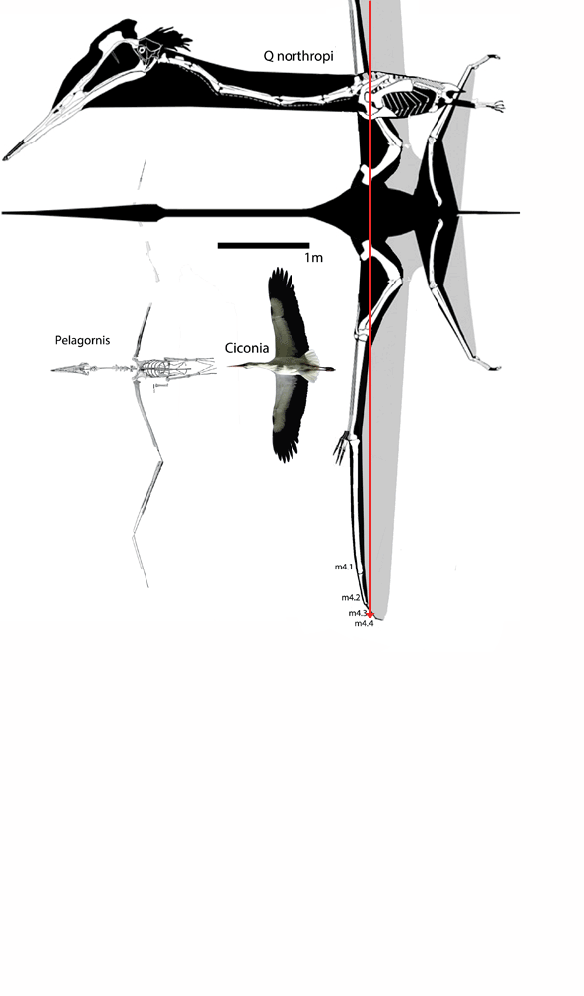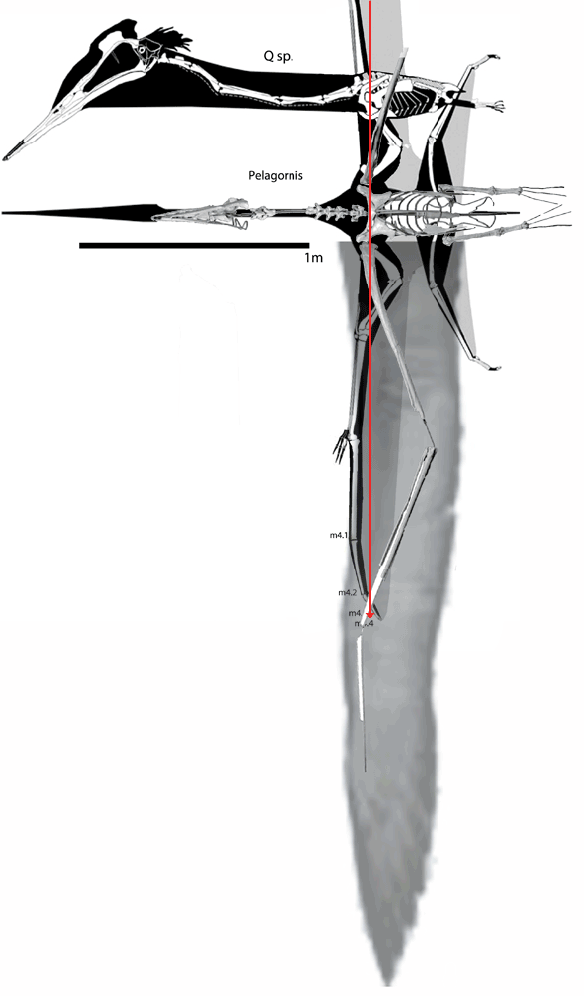
I don’t know why,
but today’s leading pterosaur experts are actively ignoring the data from the last twenty years while inventing their own fanciful versions of what pterosaurs looked like (Fig. 1) – while claiming to be the latest word on the subject. Today we’ll be looking at a short paper from the latest Flugsaurier book by Hone, Witton and Martill 2017. And we’ll criticize the artwork that crystalizes their latest intentions. This is part 1.
For some reason
Hone, Witton and Martill like to show ancient cartoons that have little to no bearing on the present knowledge base (Fig. 1). I think it’s an English thing since most, if not all of the old engravings are indeed English in origin and easily lampooned. ‘See how far we’ve come!’, they seem to be saying. Doing so only takes up space that could otherwise go to competing current versions – which they want to avoid.
We’ve seen this
earlier when English professor D. Naish preferred to criticize work that preceded (= was not included in) ReptileEvolution.com. He employed cartoons made by others, rather than artwork that was actually posted on the website to show how bad the whole website was.
Evidently
It’s what they like to do. Someday, perhaps, they’ll look in a mirror and see some of the faults I present here… using their own artwork – which will soon enough joint their ancient engravings in a drawer full of foolish ideas they can draw upon in future decades.

Figure 1. Images from Hone, Witton and Martill 2017 showing the ‘evolution’ of our concept of Dimorphodon. Artists are credited in the text. Compare the latest color version to tracings of the several skeletons in figure 2. The long tail is based on a disassociated fossil probably belonging to a campylognathoid.
In figure 1
images of Dimorphodon through time are presented from Hone, Witton and Martill 2017.
- Rev. GE Howman 1829. Probably based on the headless holotype BMNH R1034 (Fig. 2). The authors labeled this as ‘monstrous’ when ‘inaccurate’, ‘fanciful’ or ‘medieval’ would do.
- Owen 1870. Probably based on the short-skull specimen, BMNH 41212 (Fig. 2), along with the disassociated tail specimen. The authors labeled this rendition as ‘ungainly, bat-like’. Odd word choice when among all the presented illustrations it is the one most like Witton’s 2017 version (#5).
- H Seeley 1901. Probably based on the long-skull specimen, BMNH PV R 1035 (Fig. 2) In the their comment Hone, Witton and Martill report, ‘progressive interpretation of D. macronyx as an erect-limbed quadruped’, but note that a biped interpretation was also offered. They thought it best not to show that possibility.
- K Padian 1983. Probably based on the short-skull specimen, BMNH 41212 (Fig. 2). The authors report, ‘a highly active, bird-like digitigrade biped, a controversial interpretation that nevertheless symbolises the reinvention of pterosaurs in the late twentieth century.’ While there are minor issues associated with this figure (the orientation of fingers 1–3 and pedal digit 5, the over-extension of the metatarsophalangeal joint, the great length of the tail), it is the one that is most closely based on the skeleton (Fig. 2). BTW, when authors use the word, ‘controversial’ it usually means it does not fit their world view, but they have no evidence against it, nor any evidence to support their traditional hypothesis.
- M Witton 2017. Not sure which skeleton this one is based on as it appears to have been done entirely freehand from memory and imagination. The authors report, ‘Modern interpretation of D. macronyx adult and speculative juveniles reflecting contemporary interpretations of pterosaur soft tissues, muscle development and ecology.’ Ahem…we’ll run through this illustration step-by-step below.

Figure 2. Images of Dimorphodon from ReptileEvolution.com. The tail attributed to Dimorphodon is shown in figure 3.
You know, you really can’t go wrong
when you strictly adhere to the bones (Figs. 2,3), soft tissue (Peters 2002) and footprints of the most closely related taxa (Peters 2011), which were made by digitigrade and bipedal pterosaur trackmakers. Unfortunately no such citations appear in this chapter. Those are purposefully omitted.

Figur 3. Dimorphodon model by yours truly. The tail is too long based on the disassociated tail.
Witton
fell under the spell of the quad-launch hypothesis (Habib 2008), then took it one step further and made Dimorphodon a galloping hunter (Fig. 4), forsaking its wings and erect, digitigrade hind limbs (according to related ichnite makers) to hunt prey on mossy logs with backward pointing fingers. The finger unguals are again too small here.
While writing this I became aware
of Sangster 2003, a PhD thesis that evidently used computer modeling to show Dimorphodon was a quadruped. I have not seen the thesis and Ms. Sangster can no longer be found online. I wonder about these conclusions because:
- PhD theses are, by definition, the work of inexperience workers; and
- Sangster may have had to earn her PhD by succumbing to the unveiled interests of her English professors, as we’ve seen before here and here.

Figure 4. Galloping Dimorphodon by Mark Witton.
To counter the awkward, dangerous and ultimately unproductive
quad-launch scenario, I proposed the following bipedal launch animation (Fig. 5). It combines the hind limb leap with the first flap of the large wings to provide the maximum thrust at takeoff. In the Habib proposal, you don’t get that wing flap until later in the cycle – maybe too late in the cycle. The quad launch also depends on directing the force of liftoff through the fragile free fingers. They were not strong enough for that, especialy not when there is a better option available using giant muscles in the chest and pelvis. That’s why the sacrum is so strong, to act as a fulcrum on that long, heavy lever!

FIgure 5. Dimorphodon take off (with the new small tail).
So let’s get back
to Witton’s cover illustration (Fig. 6), which they tout as our contemporary view of Dimorphodon. I will note several inaccuracies (below). See figures 2 and 3 for accurate tracings.

Figure 6. Touted as the contemporary view of Dimorphodon, this Mark Witton illustration suffers from several fancies and inaccuracies.
- No Dimorphodon as this shape of skull.
- Needs a longer neck.
- Free fingers should be long and the unguals much larger.
- Wing appears to be too short with a too narrow wing tip chord.
- Witton wants to connect the trailing edge membrane from wing tip to ankle (or lateral toe), but look at the tremendous stretch in the membrane when that happens. Seems to be getting dangerously close to the narrow-at-the-elbow wing design of Zittel, Schaller and Peters, which they want to avoid.
- Ouch! This is a set of elongate toe bones with butt metatarsophalangeal joints – where Witton breaks them. This is not a calcar (a novel ossification on bat ankles which enters the uropatagium). One one side of these lateral toes the wing membrane attaches. On the other side the uroropatagium attaches. This is not shown in any fossil! Related taxa, from Langobardisaurus to Sharovipteryx, to Tanystropheus, with this same sort of elongate toe morphology, do not dislocate their bones this way. See Peters 2000 for a description that fits Rotodactylus tracks.
- No pterosaur has a uropatagium. This comes from a misinterpretation of Sordes. Pterosaur do have paired uropatagia.
- The tail is too large. On the BMNH 41212 fossil the traditionally overlooked tail is very small (Figs. 2, 7) This is in accord with related anurognathids. An unassociated tail has been attributed to Dimorphodon (Fig. 5) but it is robust and much longer. It probably belongs to a eudimorphodontid or campylognathoid. I”m surprised the tiny tail of Dimorphodon has gone unnoticed for so long. The specimen has been in English storage for over a hundred years. It was their responsibility for discovering this, but they chose instead to use their imaginations (Fig. 6).
- No tail vane is known for Dimorphodon. Tail vanes are found in pterosaurs like Campylognathoides and Rhamphorhynchus, both with a robust tail. Vestigial tails are unlikely to have had tail vanes.

FIgure 7. The tail of Dimorphodon (BMNH 41212 specimen). See figure 2 for reconstruction.
I’m asking my Engllsh colleagues
|to step up their game and become more professional. Otherwise chaps from across the pond are going to continue pointing out the flaws in their thinking. I’m not going to say their approach is not scientific (as they say about my work), but when you forsake accuracy for artistry, you’re treading very close to that line.
References
Habib MB 2008. Comparative evidence for quadrupedal launch in pterosaurs. Zitteliana B28:159-166.
Hone DWE, Witton MP and Martill DM 2017. New perspectives on pterosaur paleobiology in Hone DWE, Witton MP and Martill DM (eds) New Perspectives on Pterosaur Palaeobiology. Geological Society, London, Special Publications, 455, https://doi.org/10.1144/SP455.18
Peters D 2000. Description and Interpretation of Interphalangeal Lines in Tetrapods
Ichnos, 7: 11-41.
Peters D 2002. A New Model for the Evolution of the Pterosaur Wing – with a twist
Historical Biology 15: 277-301
Peters D 2011. A Catalog of Pterosaur Pedes for Trackmaker Identification.
Ichnos 18(2):114-141. http://dx.doi.org/10.1080/10420940.2011.573605
Sangster S 2003. The anatomy, functional morphology and systematics of Dimorphodon macronyx (Diapsida: Pterosauria)..Unpublished PhD thesis, University of Cambridge.