Bi et al. 2018 bring us a small Virginia opossum from the early Cretaceous
“Molecular estimates of the divergence of placental and marsupial mammals and their broader clades (Eutheria and Metatheria, respectively) fall primarily in the Jurassic period. Supporting these estimates, Juramaia, the oldest purported eutherian is from the early Late Jurassic (160 million years ago) of northeastern China. Sinodelphys, the oldest purported metatherianâis from the same geographic area but is 35 million years younger, from the Jehol biota. Here we report a new Jehol eutherian, Ambolestes zhoui, with a nearly complete skeleton that preserves anatomical details that are unknown from contemporaneous mammals, including the ectotympanic and hyoid apparatus. This new fossil demonstrates that Sinodelphys is a eutherian, and that postcranial differences between Sinodelphys and the Jehol eutherian Eomaia, previously thought to indicate separate invasions of a scansorial niche by eutherians and metatherians, are instead variations among the early members of the placental lineage. The oldest known metatherians are now not from eastern Asia but are 110 million years old from western North America, which produces a 50-million-year ghost lineage for Metatheria.”

Figure 1. Ambolestes tracing from Bi et al. 2018. plate and counter plate. Note the scale bar. This taxon is one third the size of the extant opossum. Apparently and oddly no unguals were preserved.
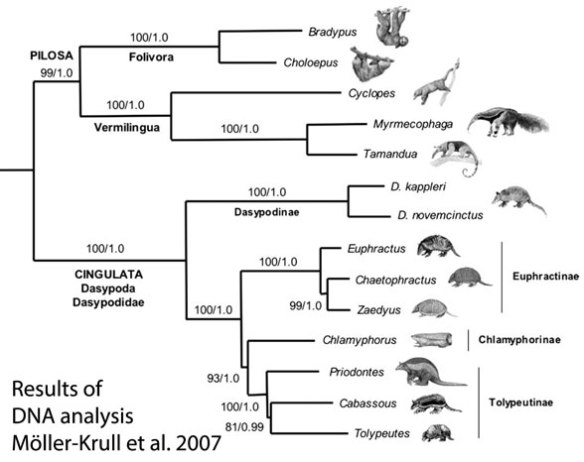
In the large reptile tree (LRT, 1131 taxa) the new fossil at the Metatheria/ Eutheria split is phylogenetically identical to and therefore congeneric with Didelphis, the Virginia opossum. So, this is pre-marsupial in a clade at the base of the marsupial/placental split. There are also a series of pre-placentals, some of which are extant and retain a reduced pouch, like Monodelphis.

Figure 2. Ambolestes skull in situ with DGS applied.
Ambolestes zhoui gen. & sp. nov. (Bi et al. 2016, 126 mya; 25cm in length) is is one third the size of the extant opossum, but otherwise nearly identical based on tested traits. In the LRT Didelphis nests with Ambolestes and they nest with Eomaia and Agilodocodon as the last common ancestors to Metatheria and Eutheria. Bi said in an interview, “Ambolestes zhoui is an early member of the placental lineage. It also carries mixed features both placentals and marsupials”. In the LRT, Ambolestes is exactly as much in the placental lineage (Fig. 4) as Didelphis is… and Didelphis has a pouch. Both have prepubic (epipubic) bones.
Since Ambulestes is congeneric with Didelphis,
you heard it hear first when the LRT nested Didelphis as the last common ancestor of Metatheria and Eutheria. Good to see confirmation.

Figure 3. Ambolestes skull reconstructed. Jaw tips restored. Lower last molar appears to be just erupting. No retroartcular process is apparent here, which sometimes happens with Didelphis (Mohamed 2018)
The Pittsburgh Post-Gazette reported,
“The well-preserved new mammal, an ancient furry creature most similar to a modern tree shrew, is named Ambolestes zhoui.” Actually Ambolestes was a little less exotic than that.
“John Wible, curator of mammals at the Carnegie Museum of Natural History, became involved in the project about two years ago. “As soon as I saw the photographs of the fossil I was like, ‘Oh my God this is amazing,’ ” he said. “It was amazingly complete. Right off the bat I saw there were skeletal parts of the body that were not known of other animals of that time period.”
But if Dr. Wible happened upon a certain type of roadkill
or went out after midnight with a flashlight he would have seen a living version of the Early Cretaceous fossil. Rather than, “this is amazing” he could have said, “we could have predicted this.”

Figure 4. Subset of the LRT focusing on basal live-bearing mammals.
The Post-Gazette also reported,
“The fossil was not allowed to leave China, said Mr. Wible, noting that this is the first paper he’s published where he’s been unable to actually hold the fossil, though he hopes to see it in person in the next few years. Instead, he relied on detailed photographs and scanned images.”

Figure 5. Didelphis, the extant opossum, a larger sister to the smaller and 126 million years older Ambolestes.
Now it’s important to remember
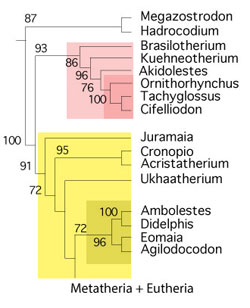
that tooth traits can converge and reverse. Think about archaeocetes (pre-whales), which have three cusps all in a row, like cynodont. Consider odontocetes, which have simple cones, like basal reptiles. Thus, tooth only taxa must be treated separately from skeletal taxa and cladograms must be based on skeletal traits, not tooth traits, which can be dangerous based on the issues that arise from the Bi et al. cladgoram (Fig. 6).
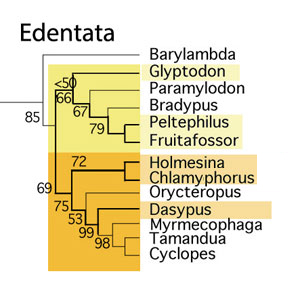
Take another look at taxa listed in Bi et al. 2018
Maelestes, when tested with more taxa, nests at the base of the tenrec/odontoete clade. Necrolestes nests with the golden mole, Chrysochloris, a basal member of Glires. Zhangeotherium is a basal pangolin. They (Bi et al. and other basal mammal workers) are going to have to expand their taxon lists to include at least all the mammals that don’t have hooves, and that includes a few that do have quasi-hooves, like the descendants of Maelestes.

FIgure 6. Cladogram published in Bi et al. 2018 with colors added to show taxa appearing elsewhere in the LRT. As you can see, this is a mess, likely created by too much emphasis on teeth traits, which converge and reverse. Treeshrew-like Maelestes, when tested with more taxa, nests at the base of the tenrec/odontoete clade. Necrolestes nests with the golden mole, Chrysochloris, a basal member of Glires. Zhangeotherium is a basal pangolin.
References
Bi S-B, Zheng X-T, X, Wang X-L, Cignetti N-E, Yang SL and Wible JR 2018.
An Early Cretaceous eutherian and the placental/marsupial dichotomy. Nature (advance online publication) DOI: https://doi.org/10.1038/s41586-018-0210-3
https://www.nature.com/articles/s41586-018-0210-3
Mohamed R. 2018. Anatomical and radiographic study on the skull and mandible of the common opossum (Didelphis marsupialis Linneaus, 1758 (in the Caribbean). Veterinary Sciences 5(44) 10 pp. doi:10.3390/vetsci5020044
https://carnegiemnh.org/press/new-mammal-fossil-provides-insights-on-early-placental-mammal-evolution/