Everyone agrees
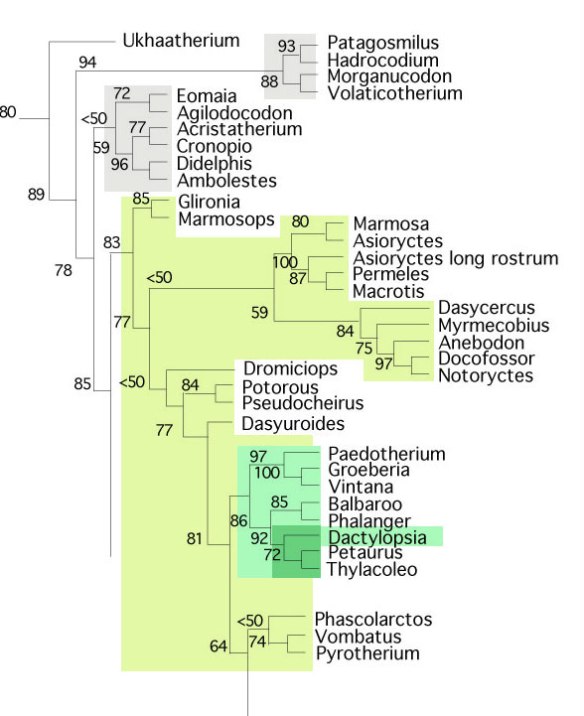
that Asiatherium (Figs, 1,2) nests close to Monodelphis, Caluromys and placentals. Trofimov and Szalay 1994 agreed. So did Denyer, Regnault and Hutchinson 2020. So did the large reptile tree (LRT, 1729+ taxa, subset Fig. 3).
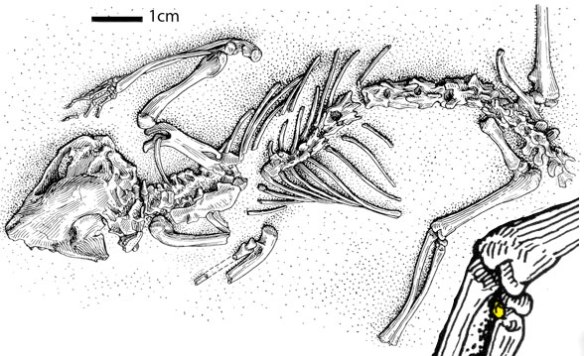
Figure 1. Asiatherium in situ from Szalay and Trofimov 1996.
Asiatherium reshetovi (Trofimov and Szalay 1994, Szalay and Trofimov 1996; PIN 3907; Late Cretaceous; 80mya; Figs. 1, 2) is a key Mongolian metathere ancestral to monodelphids and Caluromys, which is ancestral to placentals. It is derived from Triassic sisters to extant late survivors, Didelphis, Gilronia and Marmosops.

Figure 2. Asiatherium skull slightly modified (longer lateral view premaxilla to match dorsal and ventral views) from Szalay and Trofimov 1996. Colors added here.
The problem is,
according to results recovered by the LRT, mammal clade nomenclature needs to go back to basics. Several modern mammalian clade names are found to be junior synonyms of traditional clades in the LRT.
Prototheria (Gill 1872) is a junior synonym
for Monotremata (Bonaparte 1837) in the LRT.
According to Wikipedia, “Prototheria is a paraphyletic subclass to which the orders Monotremata, Morganucodonta, Docodonta, Triconodonta and Multituberculata have been assigned, although the validity of the subclass has been questioned.”
In the LRT Morganucodon is a a marsupial (see below). Docodon is a taxon within Monotremata. Triconodon is a taxon within Monotremata. Multituberculata is a clade within the placental clade Glires (Fig. 4). So, the clade Monotremata is monophyletic and has precedence.
Theria (Parker and Haswell 1897) is a junior synonym
of Marsupialia (Illiger 1811). Metatatheria (Thomas Henry Huxley 1880) is also a junior synonym of Marsupialia.
The late-surviving basalmost marsupial in the LRT (Fig. 4), Ukhaatherium (Fig. 3), has epipubic (marsupial) bones. That long rostrum indicates this taxon is close to monotremes.

Figure 3. Ukhaatherium in situ.
Unlike the monophyletic clade Monotremata,
a series of nested marsupial clades are present. The last of these gives rise to Placentalia, only one of several that lose the pouch (Fig. 4). New names are proposed here where appropriate:
- Marsupialia = Ukhaatherium and kin + all descendants (including placentals)
- Paleometatheria = Morganucodon and kin + all descendants.
- Didelphimetatheria = Eomaia and kin + all descendants
- Phytometatheria = Marmosops and kin + all descendants
- Carnimetatheria = Asiatherium and kin + all descendants
- Transmetatheria = Caluromys and kin + all descendants
- Placentalia = Vulpavus and kin + all descendants

Figure 4. Subset of the LRT cladogram of basal Mammalia. Note the new names proposed here.
Basal marsupial taxa are omnivores.
Derived phytometatheres are herbivores. Derived carnimetatheres are carnivores to hyper-carnivores. Transmetatheres (Carluromys) and basal Placentalia remain omnivores.
In the LRT Eutheria (Gill 1872) is a junior synonym
of Placentalia (Owen 1837). Omnivorous civets like Nandinia are basal placentals. Carnivora is a basal placental clade following basal placental civets.
Competing cladograms
Denyer, Regnault and Hutchinson 2020 recently looked at the marsupial patella, or more specifically the widespread absence or reduction of the kneecap. The authors concluded, “metatherians independently ossified their patellae at least three times in their evolution.”
Unfortunately, Denyer et al. tested Caenolestes, the ‘shrew opossum’. Not surprisingly it nested close to placentals in their cladogram. Caenolestes was earlier nested in the LRT within the placental clade, Glires, closer to shrews than to opossums. It has no pouch, but converges with marsupials in several aspects. Inappropriate taxon inclusion, like Caenolestes, occurs due to taxon exclusion. Excluded taxa would have attracted and removed the inappropriate taxon. Taxon exclusion plagues Denyer et al.
Historically, you may remember,
Bi et al. 2018, while presenting Early Cretaceous Ambolestes, suffered from massive taxon exclusion and traditional bias in attempting to produce a cladogram of mammals. Bi et al. recovered Sinodelphys (Early Cretaceous) and Juramaia (Late Jurassic) as ‘eutherians’. In the LRT both are monotremes.
Other basal mammal cladograms
depend too much on tooth traits. Convergence in tooth traits creates problems, as documented earlier. We’ll look at this problem in more detail soon.
The above subset of the LRT appears to be a novel hypothesis
of interrelationships. If not, please provide a citation so I can promote it.
References
Bi S, Zheng X, Wang X, Cignetti NE, Yang S, Wible JR. 2018. An Early Cretaceous eutherian and the placental marsupial dichotomy. Nature 558(7710):390395 DOI 10.1038/s41586-018-0210-3.
Denyer AL, Regnault S and Hutchinson JR 2020. Evolution of the patella and patelloid in marsupial mammals. PeerJ 8:e9760 http://doi.org/10.7717/peerj.9760
Szalay FS and Trofimov BA 1996. The Mongolian Late Cretaceous Asiatherium, and the early phylogeny and paleogeography of Metatheria. Journal of Vertebrate Paleontology 16(3):474–509.
Trofimov BA and Szalay FS 1994. New Cretaceous marsupial from Mongolia and the early radiation of Metatheria. Proceedings of the National Academy of Sciences 91:12569-12573