Updated May 15, 2023 – originally August 2019 –
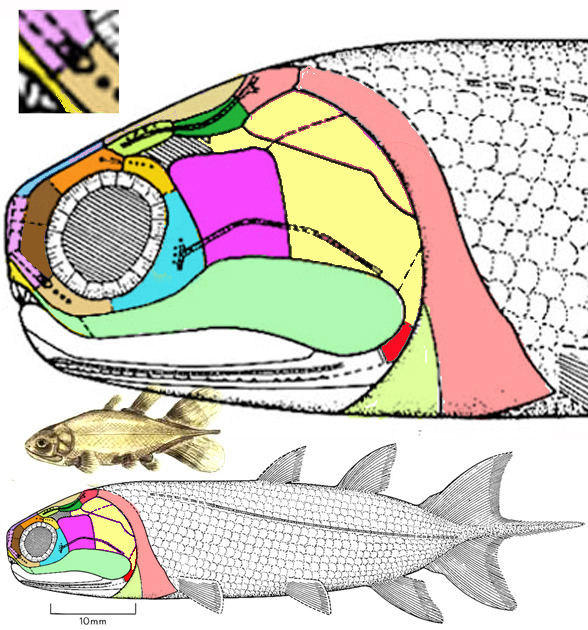
with more taxa and better scoring in the large reptile tree (LRT, 1556 taxa then, 2251 taxa today) the king mackerel, Scomberomorus (Fig 1) now nests basal to the Remora clade (Figs 1–3). This extant taxon replaces the strongly convergent cobia, Rachycentron (Fig 4), now nesting basal to barracuda in the Fundulus clade.
These examples of strong convergence are what made ray-fin fish such a difficult clade to score and understand over the past six months of focused study.

As reported at NationalGeographic.com
“The remora is so ridiculous that no one would try to make it up. The top of its head is a giant, flat suction cup. It uses the cup to lock onto the bodies of bigger animals, such as sharks, sea turtles, and whales. As the big animal swims for miles in search of a meal, the remora hangs on for the ride. When its host finds a victim, the remora detaches and feasts on the remains.”
According to Williams et al 2003, those ‘remains’ include host feces. NatGeo was using a euphemism.


As reported at NationalGeographic.com
“Their closest relatives include Mahi-Mahi and amberjacks, neither of which has anything on their head that even faintly resembles the remora’s sucker.”
The LRT does not confirm those hypothetical interrelationships. Other taxa nest closer.
For that list, see figure 1.

According to the Friedman et al 2013,
the closest relatives of remoras include the barracuda (Sphyraena), the cobia (Rachycentron, Fig 4) and early Oligocene Opisthomyzon (Figs 1,5,6).
The LRT confirms only the association of Opisthomyzon. Friedman et al did not mention or test Scomberomorus (Fig 1). In the LRT Rachycentron (Fig 4) is basal to Ductor + Sphyraena (barracuda) in the strongly convergent Fundulus clade.



As reported at NationalGeographic.com
“Britz and Johnson’s research indicates that the remora suction disk started out, improbably enough, as a dorsal fin.” (See Fig 7).
By contrast, phylogenetic bracketing in the LRT indicates the adhesion disc started out as a subdivided post-parietal (Fig 7) that expanded laterally and anteriorly during phylogeny. This is recapitulated during ontogeny as paired spines associated with the back of the skull – not associated with a single median dorsal fin associated with the spinal column.


Remoras lack a swim bladder.
Thus they are able to attach themselves in an inverted or angled position on their host (Fig 8).

Opisthomyzon glaronensis (Friedman et al. 2013; early Oligocene) is a small prehistoric remora with only a small ‘sucker’ arising from the postparietal. This specimen (Figs 5, 6) indicates that the adhesion disc originated as a subdivided postparietal, which arises from a median crest in Scomberomorus (Fig 6b).
Remora remora (Rafinesque 1810; 75cm) is the extant remora or shark-sucker. A flexible Venetian blind-like membrane rises due to blood flow atop the skull to produce suction (Flammang BE and Kenaley 2017). Hatchlings are less than a centimeter in length. At 3cm juvenile Remora has a fully formed 2mm sucking disc. By convergence with barracuda, the remora skull roof is flat and the lower jaw juts out beyond the upper one. Other traits in the LRT separate these convergent taxa.
Friedman et al report
“We demonstrate that the “interneural rays” of echeneids are homologous with the proximal‐middle radials of Morone and other teleosts and that the “intercalary bones” of sharksuckers are homologous with the distal radials of Morone and other teleosts.”
Morone is the white perch, a discoidal fresh water fish, not related to remoras in the LRT.
Echeneids are members of the remora clade.
“The “intercalary bones” or distal radials develop a pair of large wing‐like lateral extensions in echeneids, not present in this form in any other teleost.”
According to the LRT the postparietal of ray-fin fish develops phylogenetically from a series of post parietal bones that first appear in placoderms. Hiodon tergisus (Fig 10) is a more distant remora ancestor and it documents postparietal extensions and elaborations not seen in other ray-fin fish. So some strange things happen to the postparietal in this clade.
“Finally the “pectinated lamellae” are homologous with the fin spines of Morone and other acanthomorphs.”
According to Wikipedia, “A key anatomical innovation in acanthomorphs is hollow and unsegmented spines at the anterior edge of the dorsal and anal fins”. Acanthormorpha is a clade not recognized by the LRT, so this spine trait can be convergent among several clades.
Be careful of ‘key anatomical innovations’ like this, which can be convergent. Focusing on one trait is what we call “Pulling a Larry Martin.” Always nest taxa using 200+ traits from head to tail, no matter what ‘key traits’ a taxon seems to display.
“The main part of each pectinated lamella is formed by bilateral extensions of the base of the fin spine just above its proximal tip, each of which develops a row of spinous projections, or spinules, along its posterior margin. The number of rows and the number of spinules increase with size, and they become autogenous from the body of the lamellae.”
Since there is phylogenetic disagreement here due to taxon exclusion, let’s try to figure out the phylogeny first by adding taxa wherever needed. Then we can determine homology, convergence and the origin of the adhesion disc with more consensus and authority.

Finally let’s return to the Devonian to Carboniferous
and the chimaera-like extinct clade, Iniopterygidae (Fig 11). These share with remoras the trait of large elevated pectoral fins – by convergence.

Distinct from remoras,
members of the Iniopterygidae (= Iniopterygiformes, 15-46cm in length) have pectoral fins with tiny hooks. Does anyone else wonder if these elevated pectoral fins and their tiny hooks enabled iniopterygids to hitch rides on larger hosts, convergent with remoras?
This appears to be a novel hypothesis of interrelationships.
If not please provide a citation so I can promote it here.
References
Britz R and Johnson GD 2012. Ontogeny and homology of the skeletal elements that form the sucking disc of remoras (Teleostei, Echeneoidei, Echeneidae). Journal of Morphology https://doi.org/10.1002/jmor.20105 online here.
Flammang BE and Kenaley 2017. Remora cranial vein morphology and its functional implications for attachment. Scientific Reports 7(5914). https://www.nature.com/articles/s41598-017-06429-z
Friedman M, et al. 2013. An early fossil remora (Echeneoidea) reveals the evolutionary assembly of the adhesion disc. Proc. R. Soc. B 280.1766 (2013): 20131200.
Williams EH, et al. (6 co-authors) 2003. Echeneid-sirenian associations, with information on sharksucker diet. Journal of Fish Biology. 63 (5): 1176.
nationalgeographic.com/what-good-is-half-a-sucker/
scientificamerican.com/how-the-sharksucker-got-its-suction-disc/
wiki/Remora
wiki/Common_remora