
Earlier we looked at Mazin and Pouech 2020
who claimed they had discovered “the first non-pterodactyloid pterosaurian trackways.” At the time, only the abstract was available to discuss and criticize.
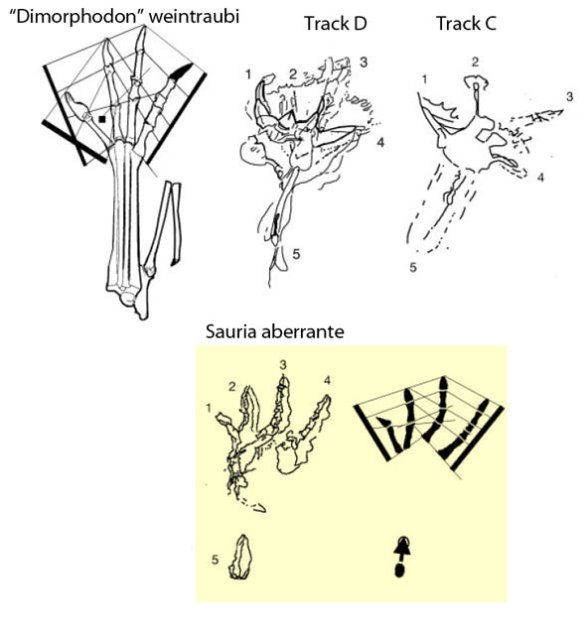
Nine years ago
Peters 2011 published anurognathid tracks, which makes them the first non-pterodactyloid pterosaurian trackways published. Notable by its exclusion, Mazin and Pouech 2020 did not cite, “A catalog of pterosaur pedes for trackmaker identification” (Peters 2011), confirming Dr. S. Christopher Bennett’s threat, “You will not be published. And if you are published, you will not be cited.”
Now that I have seen the paper and the tracks,
(Figs. 1, 2) let’s determine what sort of tetrapod made those tracks named, Rhamphichnus crayssacensis, because they don’t look like other pterosaur tracks, as workers (see below) acknowledge.
Diagnosis from Mazin and Pouech 2020:
“Quadrupedal trackway with tridactyl digitigrade manus-prints and pentadactyl plantigrade to digitigrade pes-prints. Subparallel manus digit-prints orientated anteriorly. Pentadactyl pes-prints with more or less divergent digit prints. Pedal digit V divergent and postero-laterally rejected. Manus trackway slightly to clearly wider than the pes trackway.”
Distinct from typical pterosaur manus tracks:
- tridactyl digit prints are subparallel (rather than widely splayed)
- digits are oriented anteriorly (rather than laterally to posteriorly)
- digits sometimes include additional medial and lateral impressions (never seen in other pterosaur tracks)
- no claw marks are present (that seems wrong based on Fig. 1)
- the manus impression is just anterior to the pes impression (rather than laterally and posteriorly, as in other pterosaur tracks)

Figure 1. Images from Mazin and Pouech 2020. Some manus tracks have at least four digits.
There are many
basal and derived pterosaurs with pedal digit 2 (or 2 and 3) the longest, distinct from Triassic pterosaurs. These were all examined and rejected as potential trackmakers matching Rhamphichnus for various reasons.
I also looked at 1600+ non-pterosaur trackmakers
due to the many unexpected traits (see list above) present in the Rhamphichnus tracks.
First and foremost,
the pterosaur antebrachium (radius + ulna) could not be pronated to produce anteriorly-oriented Rhamphichnus tracks. Due to folding and flying issues, pterosaurs, like birds, do not have the ability to pronate and supinate the wing. That’s why all pterosaur manus tracks are oriented laterally with fingers at full extension, impressing into the substrate. That manus digit 3 is often rotated posteriorly is a clue to its lepidosaurian ancestry. These facts form the hypothesis of a secondarily quadrupedal configuration for some, but not all pterosaurs.
One overlooked trackmaker stood out as a good match
for Rhamphichnus: the tenrec, Tenrec (Fig. 2), a small digitigrade quadrupedal mammal currently restricted to Madagascar. The medial and lateral manual digits are shorter than 2-4, which are parallel in orientation.

Figure 2. Rhamphichnus tracks compared to a Tenrec trackmaker. The brevity of pedal digit 5 is a mismatch, but a related taxon, Leptictidium, likewise reduces pedal digit 5.
One of those Tenrec sisters,
Rhynchocyon, greatly reduces manual digits 1 and 5, but pedal digit 3 is the longest.
Another Tenrec sister,
Leptictidium (Fig. 3), has a pes with a reduced pedal digit 5, but a short digit 2, but the manus is also a good match for Rhamphichnus. So there is great variation in the pes of tenrec clade members. Still, a small tenrec-like mammal remains a more parsimonious trackmaker than any Late Jurassic pterosaur. They were able to pronate the manus!

Figure 3. Elements of Leptictidium from Storch and Lister 1985.
Due to taxon exclusion,
Mazin and Pouech 2020 did not consider alternative trackmakers for the pterosaur-beach Rhamphichnus tracks that don’t match other pterosaur tracks or extremities. Now we’re stuck with an inappropriate name for these Late Jurassic tenrec tracks.
A late Jurassic tenrec?
The large reptile tree (LRT, 1637 taxa) supports the probability that a sister to Tenrec was present in in the Late Jurassic based on the coeval presence of derived members of Glires (Multiturberculata). Placental mammal fossils remain extremely rare in the Mesozoic, but these impressions add to their chronology.
It is worth repeating, due to the subject matter,
the Crayssac pterosaur beach still includes the pes of the JME-SOS 4009 specimen attributed to Rhamphorhynchus, as mentioned earlier. Here it is again (Fig. 4).

Figure 4. Crayssac track different from all others. Inset: Pes of Rhamphorhynchus muensteri JME-SOS 4009, no. 62 in the Wellnhofer catalog
twitter.com/Mark Witton mistakenly reports:
“Turns out we’ve been over-thinking it (pedal digit 5): it just lays flat on the ground during walking, like a regular toe.”
“For one, the walking fingers face forward, not sideways, as in pterodactyloids. This seems weird, but it turns out that non-ptero wing fingers fold roughly perpendicular to the walking digits.”
These basic bungles by a PhD pterosaur worker
demonstrate the dominance of myth-making among purported experts due to accepting published results like a journalist, without testing them, like a scientist. Dr. Witton’s 2013 pterosaur book is full of similar mistakes reviewed here in a seven-part series.
References
Mazin J-M and Pouech J 2020.The first non-pterodactyloid pterosaurian trackways and the terrestrial ability of non-pterodactyloid pterosaurs. Geobios 16 January 2020. PDF
Peters, D. 2011. A Catalog of Pterosaur Pedes for Trackmaker Identification
Ichnos 18(2):114-141. http://dx.doi.org/10.1080/10420940.2011.573605













 I’ve had a long history with Dr. Kevin Padian,
I’ve had a long history with Dr. Kevin Padian,











{kind=link}