
Updated August 29, 2021
with new data on Andrewsiphius: a photo of the skull (Fig. x), humerus and femur. Note the differences between the bones and the diagram (Fig. 1).

Figure x. Andrewsiphius skull
This is yet another case
of taxon exclusion and therefore… another excellent subject for a PhD dissertation!
According to Wikipedia,
“Andrewsiphius (Fig. 1) is an extinct remingtonocetid early whale known from the Eocene.”

Figure 1. The reconstructed and restored skull of Andrewsiphius from Theiwissen 2009 and colorized using DGS methods here. Tenrec skull is to scale.
Not quite.
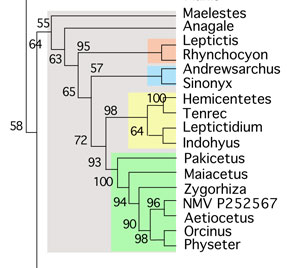
In the large reptile tree (LRT, 1695+ taxa, subset Fig. 2), which tests more taxa, Andrewsiphius nests as the proximal outgroup to Pakicetus (Fig. 3), the most primitive known walking whale. Prior studies omitted tenrecs and other basal taxa recovered by the LRT.
Andrewsiphius also nests as the proximal outgroup to extant tenrecs (Figs. 3, 5). So, if Andrewsiphius IS a walking whale, so are living tenrecs.
Andrewsiphius was derived from basal leptictids, like Leptictis. One of these leptictids is extant, the long-nosed elephant shrew, Rhynchocyon (Fig. 6), ancestor to the misunderstood giant predator, Andrewsarchus.

Figure 2. Subset of the LRT focusing on Odontoceti and their ancestors. Here is the nesting of Eocene Andrewsiphius basal to extant tenrecs between leptictids and pakicetids.

Figure Y. Taxa added August 29, 2021 and the new data from the skull photo (Fig. x) change things slightly in this subset of the LRT.
Andrewsiphius sloani (originally Protocetus sp. Salni and Mishra 1972; Thewissen and Bajpal 2009; Eocene 50mya est. 3m) was originally considered an early whale with feet, but here nests basal to smaller tenrecs, in a clade between Leptictis and Pakicetus. The rostrum was long and narrow with molars simplified and premolarized. The orbital notch is quite small, as in other tenrecs. The narrow sagittal crest is distinct from the somewhat broader crania found in living tenrecs, but similar to extinct lepitictids including Andrewsarchus.

Figure 3. Skulls of transitional taxa between tenrecs and Odontoceti (toothed whales). These include Tenrec, Lepticitidium, Pakicetus, Rhodhocetus and Orcinus.
According to Wikipedia,
“Andrewsiphius and Kutchicetus (Fig. 4) share several characteristics not present in other remingtonocetids: an elongated snout that is higher than it is wide; foramina (small holes) on the tip of the snout suggesting the presence of whiskers; eyes located dorsally near the cranial midline, resulting in an appearance of a mammalian crocodile; and a very large sagittal crest overhanging the back of the skull. Other characteristics make them distinct: the second and third upper and lower premolars are double-rooted in Andrewsiphius but single-rooted in Kutchicetus; the large diastemata in the former are absent the latter; and the tail vertebrae are more robust in Andrewsiphius.”

Figure 4. Kutchicetus, a sister to Andrewsiphius.
Molars go through a massive reversal in odontocetes and their ancestors,
from multi-rooted, multi-cusped back to single-rooted, single-cusped, as in basal reptiles.

Figure 5. Tenrec museum mount. The long snout of archaeocete and odontocete whales is already evident here.
Tenrec ecaudatus (Lacépède 1799; extant; common or tailless tenrec; 26-39cm; Fig. 5) is a Madagascar predator that gives birth to as many as 32 young and is able to hibernate for up to 9 months. Gould 1965 found evidence for echolocation in tenrecs.

Figure 6. Rhynchocyon, a living elephant shrew, is a living leptictid.
Rhynchocyon chrysopygus (Peters 1847) is the extanct golden rumped elephant shrew that nests here with the tenrecs, unlike another ‘elephant shrew’, Macroscelides, which nests with tree shrews. In Rhynchocyon the cranium is not expanded, the rotrum is long and the orbit is set posteriorly on the skull. It can run fast on narrow limbs as it spends its day seeking invertebrates in leaf litter.
Previous clades of placentals
(Carnivora, Volitantia, Primates, Glires, Scandentia) are derived from arboreal tree shrews descending from Early Jurassic sisters to arboreal Caluromys. The tenrec-odontocete clade is the most primitive clade of placentals that is terrestrial, as are all that follow except derived members of the Xenarthran (small sloths and anteaters) which exceptionally return to the trees.
I hope this stirs more interest into the tenrec origin of Odontoceti
documented in the paper “Triple Origin of Whales” available online here. It was considered a ‘just so’ story and rejected when originally submitted because it explained everything the experts had missed. The nesting of Andrewsiphius as a large Eocene tenrec supports and cements the current hypothesis of interrelationships recovered by the LRT.
If you know of any earlier iteration of this hypothesis,
let me know so I can promote it.
References
Gould E 1965. Evidence for Echolocation in the Tenrecidae of Madagascar
Proceedings of the American Philosophical Society 109 (6): 352-360. online here.
Sahni A and Mishr VP 1972. A new species of Protocetus (Cetacea) from the Middle Eocene of Kutch, western India. Palaentology 15(3):490–495.
Thewissen JGM and Bajpai S 2009. New skeletal material of Andrewsiphius and Kutchicetus, two Eocene cetaceans from India. Journal of Palaeontology 83(5):635–663.
wiki/Leptictis
wiki/Rhynchocyon
wiki/Hemicentetes
wiki/Tenrec
wiki/Andrewsiphius





























{kind=link}
{kind=link}