no doubt, a little too big to glide…
and Thylacoleo (Fig. 2) is looking even less carnivorous in phylogenetic bracketing.
Sugar gliders
(Fig. 1) are phalangers (Fig. 6), a marsupial clade nesting between kangaroos and wombats (Fig. 5).

Figure 1. Sugar glider, Petaurus breviceps, skeleton in two views, plus a skull with mandible, lacking in the skeleton.
Adding the marsupial sugar glider,
Petaurus (Figs. 1, 3), and the cuscus, Phalanger (Fig. 6), to the large reptile tree (LRT, 1231 taxa) resolves a decades-old phylogenetic problem because Petaurus, the sugar glider, nests as a sister to Thylacoleo, the marsupial lion (Figs. 2, 4). Phalanger, the cuscus, nests as their last common ancestor, which has been suggested earlier.
According to the AustraliaMuseum website
“Most palaeontologists think that the ancestors of thylacoleonids were herbivores, an unusual occurrence since most carnivores evolved from other carnivorous lineages. One proposal suggests that thylacoleonids evolved from a possum ancestor (Phalangeroidea) based on dental formula, the skull of the cuscus Phalanger, and on a phalangerid-like musculature. Alternatively, evidence from certain skull features may show that thylacoleonids branched off the vombatiform line, the lineage that includes wombats and koalas.”
In the LRT,
wombats and koalas are now sister taxa to the cuscus clade. Without the sugar glider and the cuscus, the marsupial lion earlier nested with the wombat, Vombatus.
Just to be clear,
Phalanger is not an ancestor to Didelphis, the Virginia opossum, in the LRT, even though the Australian Museum called it a ‘possum ancestor.’

Figure 2. Thylacoleo skeleton compared to Petaurus skeleton to scale.
Long thought to be a super predator,
in the midst of a clade of gentle wombat-like herbivores, Thylacoleo had, for its size, the strongest bite of any mammal, living or extinct, despite having tiny upper canines. This linking with sugar gliders further erodes the carnivorous hypothesis.

Figure 3. Skulls of the genus Petaurus with many more teeth than in Thylacoleo, but in the same general pattern. Note the lower third premolar and its similarity to the same tooth in Thylacoleo. The big organe tooth at the tip of the dentary is the canine. The lower incisors are absent.
Arboreal or not?
Wikipedia reports, “The claws [of Thylacoleo] were well-suited to securing prey and for climbing trees.” And now we know how that came to be. Petaurus, despite its arboreal abilities, does not have a divergent thumb, like the one found in Thylacoleo.
Dentary canines
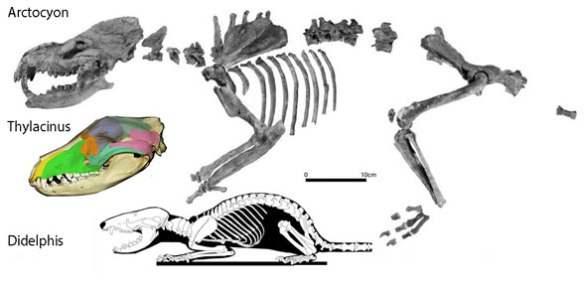
traditionally considered large, rodent-like incisors due to their placement, the anterior-most (medial-most) dentary teeth are actually canines. The incisors and their alveoli have disappeared. This can only be traced via phylogeny (see Arctocyon and Didelphis). The ancestrally small lower incisors are gone, replaced with ancestrally large large lower canines that meet medially like typical incisors. Notably, the lower canines maintain their traditional placement relationship to the upper canines (Fig. 6).
Even more interesting,
some marsupial taxa that experience a phylogenetic miniaturization, like Eurygenium (basal to Toxodon) the incisors reappear and the canines are not much larger than the incisors. That’s called a reversal or an atavism.

Figure 4. Thylacoleo skull. Many times larger than Petaurus, with fewer larger teeth, this is a giant sugar glider. The large orange tooth is the lower canine. The upper canine is a vestige.
Size
Thylacoleo was 71 cm tall at the shoulder, about 114-150cm long from head to tail tip, about the size of a jaguar.
Petaurus is 40cm long to the tail tip, about the size of a ‘flying’ squirrel. Loose folds of skin spanning the fore and hind limbs to the wrists and ankles are used to extend glides from tree to tree, or up to 140m. The diet includes sweet fruits and vegetables.

Figure 5. The sugar glider, Petaurus, in vivo. Note the wrinkled fur between the fore and hind limb. That’s the gliding membrane.
Petaurus species
According to Wikipedia, “There are six species, sugar glider, squirrel glider, mahogany glider, northern glider, yellow-bellied glider and Biak glider, and are native to Australia or New Guinea.” Whichever one is closest to Thylacoleo has not been tested or determined.

Figure 5. Subset of the LRT focusing on Marsupialia, Metatheria and then nesting of Thylacoleo.
Petaurus breviceps (Waterhouse 1839; Early Miocene to present; up to 30cm) is the extant sugar glider, a nocturnal squirrel-like marsupial able to climb trees and glide with furry membranes between the fore and hind limbs. An opposable toe is present on each hind foot. Sharp claws tip every digit.
Phalanger orientalis (Pallas 1766; 34 cm in length) is a nocturnal arboreal folivore marsupial known as thte Northern common cuscus. Commonly considered a ‘possum’ the cuscus nests between wombats and kangaroos, basal to sugar gliders and marsupial lions.

Figure 6. The cuscus (genus: Phalanger orientalis) nests with Petaurus and Thylacoleo in the LRT. Those anterior dentary teeth look like incisors, but phylogenetically are actually canines.
Thylacoleo carnifex (Owen 1859; Pliocene-Pleistocene; 1.14 m long) was a giant sugar glider like Petaurus. Thylacoleo had the strongest bite of any mammal with the largest, sharpest molars of any mammal. It had fewer but larger teeth than Petaurus. The manus included retractable claws. The pes had a very large heel bone (calcaneum). This supposedly carnivorous ‘marsupial lion’ nests with herbivores. Pedal digit 1 likely had a phalanx and claw, but it has not been shown.
References
Goldingay RL 1989. The behavioral ecology of the gliding marsupial, Petaurus australis. Research Online. University of Wollongong Thesis Collection. PDF
Owen R 1859. On the fossil mammals of Australia. Part II. Description of a mutilated skull of the large marsupial carnivore (Thylacoleo carnifex Owen), from a calcareous conglomerate stratum, eighty miles S. W. of Melbourne, Victoria. Philosophical Transactions of the Royal Society 149, 309-322.
Waterhouse GR 1838. Observations on certain modifications observed in the dentition of the Flying Opossums (the genus Petaurus of authors). Proceedings of the Zoological Society of London. 4: 149–153.
wiki/Petaurus
wiki/Thylacoleo
https://australianmuseum.net.au/thylacoleo-carnifex