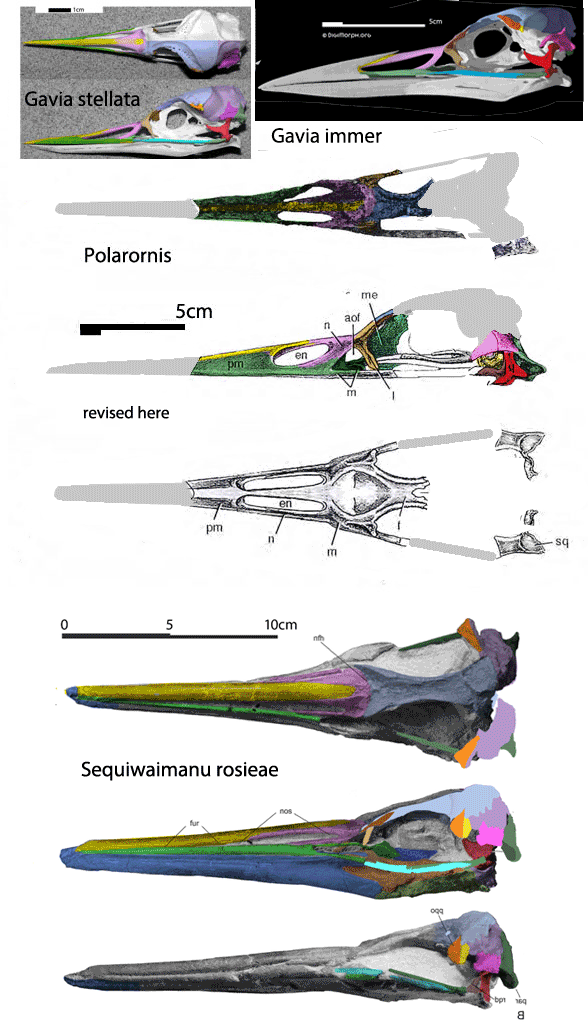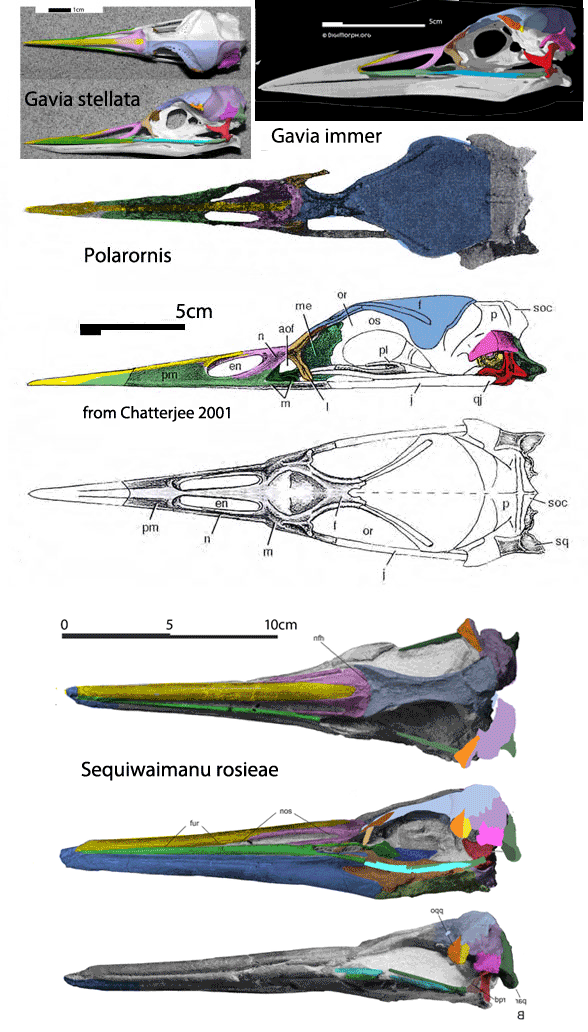
Before the genesis of the LRT
whale workers correctly took whale ancestry back to the newly discovered ‘walking whales’, Pakicetus and Indohyus (Figs. 1, 2).
Then, naturally enough, whale workers wanted to know
which taxa gave rise to ‘walking whales’. After Gingerich et al. 1994 and 2001 reported that Artiocetus and Rhodhocetus had artiodactyl-like ankles, Thewissen et al. 2007, O’Leary and Gatesy 2008 and Spaulding et al. 2009 added extant artiodactyls (Fig. 2) to their cladograms.
While adding taxa is usually a good idea
all three teams cherry-picked irrelevant taxa (Fig. 2) and omitted relevant taxa (Figs. 1, 4). As the LRT documents, sometimes it is better to simply add as many taxa as possible and let the software tell you which taxa are relevant.

By adding previously omitted taxa
the large reptile tree (LRT, 2041+ taxa) recovered walking whale ancestors that look much more like pakicetids (Fig. 1). Previously omitted tenrecs and elephant shrews look much more like pakicetids than cattle, pigs or deer do. And these non-artiodactyls have artiodactyl-like ankles.
The LRT also recovered
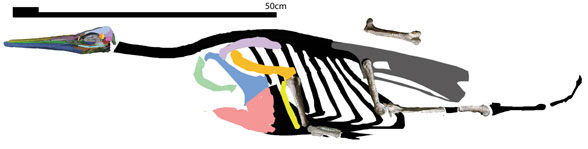
several taxa between Mesonyx and mysticetes (Fig. 4) and… big surprise… they were totally unrelated to pakicetids and odontocetes.
The splitting of the traditional clade ‘Cetacea’
happened here several years ago (October, 2016).
Decades earlier
Van Valen 1968 discussed this possibility based on a long list of physical differences. It turns out Van Valen was correct. Myticetes and odontocetes are not related to one another in the LRT. Whale workers still haven’t figured that out. According to university textbooks, and recent manuscript reviews (see below), academics are still attached to dissimilar artiodactyls, none of which show any affinity to swimming or water as a niche environment for finding their food.

Simply adding more taxa
moves artiodactyls away from Pakicetus and Indohyus and moves previously omitted tenrecs and elephant shrews closer to these ‘walking whales’. The extant elephant shrew, Rhynchocyon (Fig. 1) also has artiodactyl-like ankles, but no one noticed that until the LRT shed light on this taxon. Taxon exclusion is an easy sin to commit because it’s a sin of omission. You can’t understand and incorporate what you ignore or are not aware of.
Some of the taxa related to Pakicetus recovered by the LRT
(e.g. Rhynchocyon, Hemicentetes, Tenrec, Fig. 1) are still alive today and therefore should garner more attention in odontocete origin studies. For instance Tenrec finds prey by echolocation (Gould 1965), lacks a scrotum and travels in pods.
Unfortunately
whale workers and university textbook writers still prefer their whales to stay monophyletic and for all whales to evolve from unspecified artiodactyls. Now, even though we know better, we shouldn’t expect any changes in textbooks for another several decades. Paleontology accepts discoveries much more slowly than any other science, as Yale professor John Ostrom lamented before his passing.
Also in the LRT mix:
giant Andrewsarchus was also recovered close to the elephant shrew, Rhynchocyon (Fig. 1). Spaulding et al. 2009 (Fig. 2) included Andrewsarchus in whale origin studies, but did not understand ‘the big picture’ recovered by the LRT. They mistakenly considered Andrewsarchus another artiodactyl due to taxon exclusion.

Mysticete (baleen whale) ancestors
are an entirely distinct lineage (Fig. 3), as we learned earlier here. Now we understand the various gradual stages of mysticete evolution very well. And by ‘we’ I mean Pterosaur Heresies readers. Whale workers at the university level are still several years behind on this subject (Fig. 2).

Whale workers mistakenly put their focus on artiodactyls,
following Gingerich et al. 1994 and 2001. They should have expanded their taxon lists to include desmostylians, tenrecs and elephant shrews. Their misplaced results (Fig. 2) failed to document the gradual accumulation of traits recovered in the LRT for odontocete ancestors (Fig. 1) and mysticete ancestors (Fig. 4). It is disappointing that none of the academics recognized the folly of their hypotheses. Each team was guilty of “Pulling a Larry Martin” by putting all their faith in one cherry-picked trait, the ankle shape, rather than letting software consider the sum of every trait for a wider gamut of included taxa.
Sadly,
when walking whale expert Phil Gingerich refereed the manuscript for “The Triple Origin of Whales“, he chose to reject it, calling it a “Just So Story”. Given that Gingerich became famous for discovering walking whales and associating them with artiodactyls, perhaps he didn’t want that fame tarnished by admitting his team and his colleagues made a mistake exposed by an amateur armed with nothing more than a wider gamut taxon list. That sin is all too common at the university level and it’s a sin of commission. We’ve seen this ‘rather not know‘ attitude from professors dozens of times in paleontology. That’s never good for science. If a hypothesis is wrong, show why it is wrong. Don’t ignore it, suppress it or call the author a crank or pseudoscientist.
Happily,
if you’re interested in whale origins, you can join hundreds of researchers who have already downloaded and read the illustrated manuscript for “The Triple Origin of Whales” online here at ResearchGate.org. If you’re interested in odontocete and mysticete ancestors back to Ediacaran worms, visit the large reptile tree.
References
Gingerich PD, et al. 1994. New whale from the Eocene of Pakistan and the origin of cetacean swimming. Nature 368 (6474): 844–847.
Gingerich, PD et al. 2001. Origin of whales from early artiodactyls: hands and feet of Eocene Protocetidae from Pakistan. Science 293 (5538): 2239–2242.
Gould E 1965. Evidence for Echolocation in the Tenrecidae of Madagascar
Proceedings of the American Philosophical Society 109 (6): 352-360. online here.
Huggenberger S, Leidenbere S and Oelschläger HHA 2018. Asymmetry of the nasofacial skull in toothed whales (Odontoceti). Journal of Zoology DOI: 10.1111/jzo.12425
O’Leary MA and Gatesy J. 2008. Impact of increased character sampling on the phylogeny of Cetartiodactyla (Mammalia): Combined analysis including fossils. Cladistics 24:397–442.
Peters D unpublished. The triple origin of whales. PDF
Spaulding M, O’Leary MA and Gatesy J 2009. “Relationships of Cetacea (Artiodactyla) among mammals: increased taxon sampling alters interpretations of key fossils and character evolution”. PLOS ONE. 4 (9): e7062. doi:10.1371/journal.pone.0007062
Thewissen JGM, Cooper LN, Clementz MT, Bajpai S and Tiwari BN 2007. Whales originated from aquatic artiodactyls in the Eocene epoch of India. Nature 450:1190–1195.
Van Valen L 1968. Monophyly or diphyly in the origin of whales. Evolution. 22 (1):37–41.
wiki/Rodhocetus
wiki/Artiocetus