Venturing into invertebrates again today. This blogpost follows another posted a few days ago comparing the Early Cambrian trilobite with a telson, Pseudosaukianda (Fig 1), to the extant horseshoe crab (Limulus, also with a telson. As before, colors replace labels, to make homologous structures easier to see, scan and understand. As before, trilobites evolved from flatworm and anomalocarid ancestors.
Today the Silurian sea scorpion, Eurypterus (Fig 1),is compared to the extant terrestrial scorpion, Apistobuthus(Fig 1), and to Pseudosaukianda and Limulus (Fig 1). No phylogenetic analysis is attempted here. This is just a preliminary comparison of traits that are shared, absent and/or modified.
Evolving traits include cephalon size, chelicerae (red limb) size, oral leg length and abdomen reduction. After Pseudokianda the thorax and associated limbs are absent so oral legs are used for locomotion. Oral limb number is conserved.
Figure 1. Graphic comparisons of the Silurian sea scorpion, Eurypterus, the extant land scorpion, Apistobuthus, the extant horseshoe crab, Limulus, the Early Cambrian trilobite, Pseudosaukianda and others. Not to scale. Colors indicate homologous structures. The mouth (oral cavity) lies medial to the oral limbs, ventral to the animal, a primitive trait going back to trilobite and flatworm ancestors.
Loss of antennae is a shared trait (unknown in Pseudosaukianda Fig 1).
Respiration takes place posterior to the limbs in book lungs, which become increasingly internal and fed by tiny holes (spiracles) in spiders and scorpions. Extant sea spiders virtually lack an abdomen (a vestige remains), but have a high surface-to volume ratio to oxygenate internal tissues.
Feeding takes place medial to the oral limbs. As in their flatworm and anomalocarid ancestors, (Fig 2) spiders vomit digestive fluid over prey held ventrally beneath the oral cavity between the elbows of the oral limbs transformed into walking legs.
Eyes remain dorsal to the cephalon in all taxa.
Following the tenets of monophyly, since spiders and scorpions are also extant descendants of trilobites with a telson (Fig 1), this clade of trilobites survived to the present day by evolving and radiating into new forms while keeping a basal bauplan. This is contra traditional thinking about trilobite extinction.
Figure 2. When a planula evolves a ventral opening the internal cells can spill out to cover prey items, the retreat again to continue digestion, as in flatworms. Spiders still do this. Trilobites and anomalocarids, with their central ventral mouth parts (= toothed limb elbows) likely did the same.
Interesting that the practice of external digestion in spiders is retained from flatworms (Fig 2). That means (via phylogenetic bracketing) transitional taxa, like anomalocarids and trilobites practiced external digestion, too. That solves so many problems and overturns a myth or two.
This appears to be a novel hypothesis of interrelationships and feeding practices. If not, please provide a citation so I can promote it here.
Aria and Caron 2017 described a new Burgess Shale fossil with a bivalve carapace and crustacean-like pincers, Tokummia (Fig 1 top). The authors considered it the oldest member of the clade Mandibulata (= myriapods, spiders, scorpions, crustaceans and insects). Tokummia may be the oldest member of this clade, but is it the most primitive?
Apparently a common crawdad (Fig 1) is more primitive, closer to trilobites that have more limb-like mandible parts, more limb-like limbs and no bivalve shell.
Figure 1. Taxa in the evolution of Tokummia from trilobites with a slender telson. Here Emeraldella is the first to have uropoda (posteriormost flipper-like appendages beneath the telson. Colors represent homologous parts. Note how easy it is the ‘read’ how evolution adds/substracts limbs, and moves the oral activity surrounded by chewing limbs further forward through time, ultimately becoming a mouth surrounded by small limbs transformed into mandibles, etc.
Tokummia is apparently close to extant tiny clam shrimp (= chonchostracan), which also have pincers, but they bring prey into their shells by waving their antennae creating a current into the pincers and mouth parts. The two-piece hinged carapace draped over the thorax + limbs is a derived trait. 50+ limbs and gill pairs is also a derived trait. Therefore, shrimp-grade forms without pincers likely preceded Middle Cambrian Tokummia.
Pre-pincer crustacean taxa like Hutchinsoniella and Heterophlias (Fig 1) are likely more primitive than shrimp-grade crustaceans. There’s no phylogenetic analysis here. Too few taxa are involved. At this early stage in the scientific method, gathering data, emerging patterns start to become apparent, subject to later changes.
From the Aria and Caron abstract: “Retracing the evolutionary history of arthropods has been one of the greatest challenges in biology. During the past decade, phylogenetic analyses of morphological and molecular data have coalesced towards the conclusion that Mandibulata, the most diverse and abundant group of animals, is a distinct clade from Chelicerata, in that its members possess post-oral head appendages specialized for food processing, notably the mandible. The origin of the mandibulate body plan, however, which encompasses myriapods, crustaceans and hexapods, has remained poorly documented.”
“Here we show that Tokummia katalepsis gen. et sp. nov., a large bivalved arthropod from the 508 million-year-old Marble Canyon fossil deposit (Burgess Shale, British Columbia), has unequivocal mandibulate synapomorphies, including mandibles and maxillipeds, as well as characters typically found in crustaceans, such as enditic, subdivided basipods and ring-shaped trunk segments.”
Note the reduction in number and size from six long oral elements/limbs/gills, as in trilobites, to two short oral elements comprising the derived mandibles in Tokummia.
Wikipedia reports, “Tokummia katalepsis is a fossil arthropod from the Burgess Shale as found in a quarry in Marble Canyon in Canada. The fossil is 508 million years old. At the front of the animal are mandibles with pincers. This is the oldest fossil with pincers. It has more than 50 pairs of legs. The animal is about 10 cm long and has a two piece carapace on its back. It was a bottom feeder, being able to walk on the sea floor, and to occasionally swim. The animal legs have endites which are small spikes on the legs. It has one pair of antennae.”
The discoverers claim that Tokummia along with Branchiocaris, Canadaspis and Odaraia are very early crustaceans ancestral to the Mandibulata grouping, which includes millipedes, insects and crustaceans.
Colors seem to help identify homologous structures in diagrams (Fig 1) more simply, clearly and completely than traditional dashed lines leading to traditional abbreviations on traditional line art with a traditional key to those abbreviations traditionally located at the bottom of the graphic in traditional books. I hope colors work as well for you in your own studies. Ultimately labels must be applied, but colors will do for now.
As reported earlier, most trilobite clades became extinct. The clade with a telson survived, evolving to become horsehoe crabs, crustaceans (see above) and other extant taxa we’ll look at over the next few days.
References Aria C and Caron J-B 2017. Burgess Shale fossils illustrate the origin of the mandibulate body plan. Nature 545(7652): DOI:10.1038/nature22080
Martinez et al. 2022 described an unusual (for its stratum) dentary tip from Triassic Argentina (Fig 1). It had widely spaced teeth, separated by deep concavities, and the mandibles were fused along the mid-line (Fig 1). The authors named the jaw tip, Pachagnathus benitoi (PVSJ 1080, Fig 1) and nested it with the coeval, but dissimilar pterosaur, Raeticodactylus (Figs 2a,b), from Switzerland.
If only Pachagnathus was found in Early Cretaceous strata, it would have been easily and readily identified as a typical ornithocheirid (Figs 3, 4). Conspicuous by absence, ornithocheirids were omitted from the cladogram of Martinez et al. 2022 (Fig 5).
Figure 1. What little is known of Pachagnathus benitoi. The rest of the mandible is restored based on coeval, but geographically and phylogenetically distant Rhaeticodactylus (Fig 2). The published anterior – posterior orientation might need a switch. Hard to tell here without the usual ‘landmarks’. More importantly, compare this scrap to Early Cretaceous ornithocheirds with strongly similar traits in figures 3 and 4.
The skull of Raeticodactylus was preserved flattened in left lateral view (Fig 2a), so the width of the dentary was not apparent. Rotated palatal elements permit a typical (= not narrow) reconstruction of the skull (Fig 2b).
Figure 2a. Snout of Raeticodactylus. This morphology of the dentary does not match that of Pachagnathus.
The dentary of Raeticodactylus (Fig 2a) lacks widely spaced teeth separated by fossae/ concavities/ indentations, as seen in Pachagnathus. Rather the dentary teeth of Raeticodactylus are in contact with one another.
Figure 2b. Raeticodactylus reconstructed. Note the slender bones in the humerus, unmatched by any other pterosaur. The width of the skull was based on palatal elements. The width of the unknown skull of Pachagnathus was likely much narrower based on the narrow anterior mandibles in figure 1.
Other toothed pterosaur dentaries don’t become so narrow, indented between the teeth and medially fused like Pachagnathus (Fig 1) until the appearance of Early Cretaceous ornithocheirids (Figs 3, 4). That’s what makes Late Triassic Pachagnathus appear to be so derived and anachronistic (= not of its time).
Figure 3. Early Cretaceous Targaryendraco mandible tip showing widely spaced teeth on fused mandibles. Concavities between alveoli are shallower here than in Pachagnathus in figure 1. No ornithocheirds are shown in the cladogram of Martinez et al. 2022 shown in figure 3.
According to Wikipedia, “PVSJ 1080 was discovered in a horizon of reddish muddy sandstone just 30 m below the top of the formation, beneath the unconformably overlying Cretaceous aged El Gigante Group.”
No Jurassic strata separate the Triassic and Cretaceous strata where Pachagnathus was found. Is it possible that a Cretaceous scrap of fossil fell out of cliff to a new resting place in Triassic strata at some time in the past 100 million years? If Pachagnathus was reworked, that would help explain the presence of a typical ornithocheirid-like set of narrow fused dentaries in the Late Triassic when all other Triassic pterosaurs had ordinary dentaries (Fig 2a).
Those dentary indentations in Early Cretaceous ornithocheirids permit the upper teeth to slip past the lower teeth while keeping the rostrum extremely narrow (Fig 4) as in Pachagnathus.
Figure 4. Early Cretaceous Ferrodraco from Australia. Note how the upper teeth insert into the dentary concavities on a narrow indented dentary with widely spaced teeth, as in Pachagnathus.
According to Wikipedia describing Pachagnathus, “The symphysis is notably long, including at least five pairs of teeth and likely to have been even longer. This symphysis is very narrow and laterally compressed, so much so that the edges of the jaw tips are parallel to each other, with a preserved length of 61.5 mm but a maximum width of only 12.2 mm.”
That’s odd because only one pair of teeth is illustrated (Fig 1) and the other two illustrated teeth are presented as unpaired.
According to Wikipedia, and citing Martinez et al. 2022. “Such a combination of midline keel and a series of fossae is not known in any other pterosaur.”
Except in Targaryendracoand its ornithocheirid kin (Figs 3, 4), both from the Early Cretaceous where this keel and fossae morphology is typical. The authors should have reported this morphology is not known in any other Triassic pterosaur.
Too little is known of Pachagnathus to add it to the large pterosaur tree (LPT, 262 taxa), but what little is known of it’s morphology clearly seems to be ornithocheirid.
From the Martinez et al. 2022 abstract: “Pterosaurs were the first vertebrates to evolve powered flight. The timing of their origin is still debated, and hypotheses range from the end of the Permian Period, to the lower Mesozoic Era, and through to the Middle–Late Triassic epochs.”
I haven’t seen any evidence or speculation for a Permian origin for pterosaurs. Please provide a title and citation if you have seen this.
To set the record straight (again) Middle TriassicCosesaurus and its Late Triassic kin are valid and verified pterosaur precursors, now known for 22 years (Peters 2000). It remains odd that pterosaur workers won’t consider these taxa or test these taxa, but they might cite, then dismiss Peters 2000 in their misguided attempts at finding pterosaur ancestors within the Archosauriformes.
“Regardless of when they originated, the oldest records are restricted to the Upper Triassic Norian Stage in the northern hemisphere (Europe, USA and Greenland).”
‘Regardless‘,apparently means the authors think they have a derived Triassic pterosaur, regardless of how or when pterosaurs came to be, a subject they are both interested in and detach themselves from.
Outgroups are important. Unfortunately the cladogram by Martinez et al. 2022 (Fig. 3) lacks outgroups. That includes the Triassic pterosaur, Bergamodactylus, which nests as the most primitive pterosaur in the LPT.
“We report two new raeticodactylid pterosaurs, Yelaphomte praderioi gen. et sp. nov. and Pachagnathus benitoi gen. et sp. nov. from the upper Norian to Rhaetian Quebrada del Barro Formation in north-western Argentina. The new specimens (an isolated dentary symphysis, partial rostrum, and distal half of ulna) are the first unequivocal Triassic records of pterosaurs in the southern hemisphere, confirming that the absence of pterosaurs outside north-western Pangaea during the Late Triassic was the result of poor sampling rather than true absence.”
Excellent! And glad to see this. Now that the former Northern Hemisphere paradigm is broken, we can hopefully and eagerly be looking for more Triassic pterosaurs from Argentina and elsewhere below the Equator. Or not.
At the same time, let’s encourage Martinez et al. to put those omitted Early Cretaceous ornithocheirids back into their cladogram (Fig 5) to see if Pachagnathus (Fig 1) is attracted to ornithocheirids. Or not.
Figure 5. Cladogram from Martinez et al. 2022. These workers employ many taxa based on scraps while omitting many taxa based on complete skeletons. That produces odd results. Nesting anurognathids with Pterodactylus is odd. Kryptodraken is Sericipterus. Ornithocheirids were omitted from this cladogram.
With pterosaurs, as bizarre as they are, anything can happen. Even so, we can all wish some ornithocheirids with fused, indented dentaries sporting widely spaced teeth had not been omitted from the Martinez et al. cladogram (Fig 5). Given the morphology of Pachagnathus, the omission of ornithocheirids is conspicuous, not innocuous and certainly not warranted. A genuine pterosaur dentary from the Triassic of Argentina is a headline. A reworked fossil from the Early Cretaceous is not. By putting omitted ornithocheirids back into their cladogram the authors would have been able to confirm or refute the possibility of a reworked and no longer bizarre fossil prior to publication and publicity.
You can learn more about Brian Andres and his years of struggles with pterosaur morphology and phylogeny due to taxon exclusion by clicking here. Several links to prior blogposts will appear. One link is included below.
References Martínez RN, Andres B, Apaldetti C and Cerda IA 2022. The dawn of the flying reptiles: first Triassic record in the southern hemisphere Papers in Palaeontology 8(2): e1424 doi: https://doi.org/10.1002/spp2.1424 Peters D 2000b. A Redescription of Four Prolacertiform Genera and Implications for Pterosaur Phylogenesis. Rivista Italiana di Paleontologia e Stratigrafia 106 (3): 293–336.
Figure 1. Cast of the UNSM 93000 specimen of Nyctosaurus. Missing parts are modeled here.
Freehand illustrations too often reflect the paradigms and wishes of the artist.
By contrast manipulable bone-by-bone sculptures (Fig 2) based on nearly complete specimens (Fig 1) minimize academic and artistic preconceptions and myths.
Figure 2. Wood and wire Nyctosaurus sculpture based on UNSM 93000 (figure 1).
Adding a wing membrane to a bipedally standing graphic of Nyctosaurus (Figs 2, 3) demonstrates how the only membrane shape preserved in pterosaurs, the shallow chord stretched between the elbow and wing tip (Zittel 1882, Peters 1995, 2002, 2009), works well. This bauplan permits complete folding of the membrane when terrestrial and plenty of wing chord when airborne.
Figure 3. Nyctosaurus in dorsal view. Wings open and close every ten seconds or so. The short chord membrane shown here is the only one preserved in all pterosaurs. and is the only one that folds away virtually completely as shown here. Pterosaur precursors start the membrane distally. The bat-wing paradigm is a myth supported by PhDs who prefer textbook traditions to precision.
Nyctosaurus sp. UNSM 93000 Brown 1978, 1986, Figs 1–4) was derived from a sister to Nyctosaurus gracilis and phylogenetically preceded the crested Nyctosaurus specimens. Except for the rostral tip, the skull and distal cervicals are missing. This specimen fuses two wing phalanges resulting in only three wing finger bones. Note the free fingers are reduced to vestiges without claws.
Figure 4. Nyctosaurus sp. UNSM 93000 in lateral view and in dorsal view. Like to see Nyctosaurus perform a quad-launch doing a push-off with those hind limbs and a push-up with its folded wings? That would lead to a charge of mistreatment of animals. So much better to stand bipedally (Fig 2), open those wings at leisure (Fig 3) then take off with both trust and lift from a single coordinated downstroke + leap.
Please share this post with your pterosaur artist and expert friends so they will discontinue the myth of the bat wing pterosaur. I realize this is a naive request, but letting the PhDs and artists get away with their inept tracings and wishful ‘shrinking’ is not good science.
Figure 5. Problems with the Elgin, Hone and Frey (2011) pterosaur wing model with corrections proposed by Peters (2002). Lower right figure shows how they even cheated the graphics from Peters 2002 to make it look untenable when they could have just used an illustration as is. Disappointing that the professionals keep acting so poorly. Click to enlarge.
The Vienna Pterodactylus specimen (Fig 6) was illustrated freehand by Elgin, Hone and Frey 2011 (Fig 5 top), but given imaginary deep chord bat wings (Fig 5). In their minds the membranes were ‘suffering from shrinkage’ (yes, they used the word ‘shrinkage‘) due to their ingrained paradigms, wishful thinking and shoddy tracings. Plus they had a wish to not confirm the earlier work of Peters 2002. If this sort of cheating is what they teach and condone at the university level, and if this is how paleo students get a good grade, I am so glad I never went that route. Peer-group pressure must be a powerful force among academics.
Figure 6. The Vienna specimen of Pterodactylus preserves wing membranes in the folded position. Here animation opens the wings. The same specimen was illustrated by Elgin, Hone and Frey 2011 but given deep chord bat wings (Fig 5) in their minds ‘suffering from shrinkage’ due to their ingrained paradigms, wishful thinking and shoddy tracings. Plus their wish to not confirm the earlier Peters 2002. If this sort of cheating is what they teach and condone at the university level, and if this is how you get a good grade, I am so glad I never went that route.
In the words of Eric “Otter” Stratton (in the 1978 comedy, Animal House: “Now we could do it with conventional weapons, but that could take years and cost millions of lives. No, I think we have to go all out. I think that this situation absolutely requires a really futile and stupid gesture be done on somebody’s part!”
References Brown GW 1978. Preliminary report on an articulated specimen of Pteranodon Nyctosaurus) gracilis. Proceedings of the Nebraska Academy of Science 88: 39. Brown GW 1986. Reassessment of Nyctosaurus: new wings for an old pterosaur. Proceedings of the Nebraska Academy of Science 96: 47. Elgin RA, Hone DWE and Frey E 2011. The extent of the pterosaur flight membrane. Acta Palaeontologica Polonica 56 (1), 2011: 99-111. doi: 10.4202/app.2009.0145 Peters D 1995. Wing shape in pterosaurs. Nature 374, 315-316. Peters D 2002. A New Model for the Evolution of the Pterosaur Wing – with a twist. – Historical Biology 15: 277–301. Peters D 2009. A reinterpretation of pteroid articulation in pterosaurs. Journal of Vertebrate Paleontology 29:1327-1330. Zittel KA 1882. Über Flugsaurier aus dem lithographischen Schiefer Bayerns. Palaeontographica 29: 7-80.
Müller and Garcia 2022 report on their discovery of a right femur from the Middle Triassic (Fig 1). Their headline calls it the “oldest dinosauromorph from South America” and their highlights report it fills “a biogeographical gap”. The abstract then qualifies the claims of their headline as they report, “the new specimen is potentially the oldest dinosauromorph from South America, narrowing the biogeographical gap between Africa and Argentina during the early radiation of dinosauromorphs.”
Figure 1. Oldest dinosauromorph femur from Müller and Garcia 2022. PVL 4597 added here.
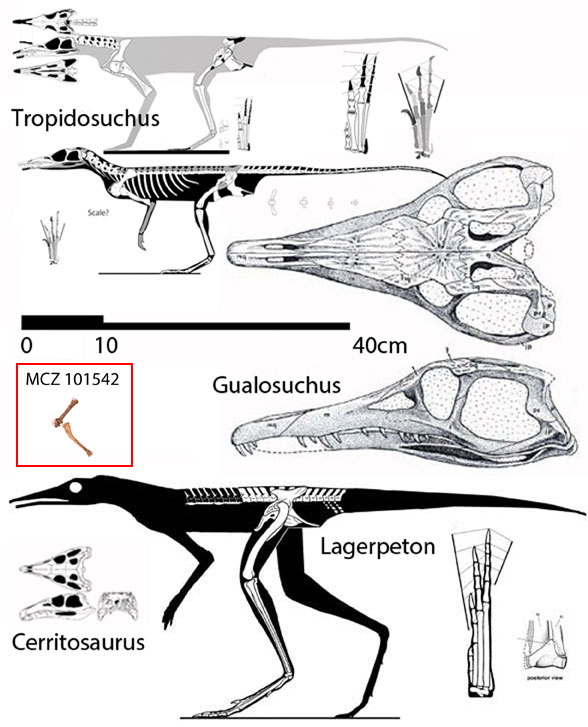
Unfortunately Müller and Garcia don’t realize there is no such thing as a ‘dinosauromorph‘. That’s an invalid clade commonly associated with lagerpetetids, a clade of bipedal proterochampsids close to Tropidosuchus(Fig 2, Novas and Agnolin 2016, but you heard it here first in 2011), therefore not related to dinosaurs. Adding taxa reveals the hypothesis that lagerpetids are convergent with dinosaurs in their bipedal abilities, not homologous.
Figure 2. Tropidosuchus and Lagerpeton compared to the new material (MCZ 101542).
According to the large reptile tree (LRT, 2061 taxa) dinosaurs and crocodiles share a last common ancestor just prior to their phylogenetic split. In the LRT that last common ancestor is the Tucuman specimen (PVL 4597, Fig 1 top) a basalmost archosaur often wrongly attributed to Gracilisuchus.
Only Crocodilomorpha and Dinosauria comprise the Archosauria (by definition and accoding to the LRT). All other Triassic reptiles (e.g. Poposauria, Aetosauria, Rauisuchia) are outgroups. Lagerpeton, Tropidosuchus and related proterochampsids arise from a clade that includes phytosaurs, choristoderes and proterosuchids, which split from the Euparkeria clade some time earlier in the Late Permian. Taxon exclusion, university textbooks and out-of-date traditions have kept certain myths alive to be taught to the next generation of tuition-paying students.
Figure 3. The origin of dinosaurs in the LRT to scale. Gray arrows show the direction of evolution. This image includes Decuriasuchus, Turfanosuchus, Gracilisuchus, Lewisuchus, Pseudhesperosuchus, Trialestes, Herrerasaurus, Tawa and Eoraptor. Note the phylogenetic miniaturization at the origin of Archosauria (Crocs + Dinos).
A previous paper by Müller 2022 eagerly followed Ezcurra et al 2020 who also hoped lagerpetids were close to pterosaurs. Taxon exclusion was also a problem in both papers. Müller and Garcia, like other current authors, are omitting basal bipedal crocodylomorphs and proterochampsids like Tropidosuchus (Fig 2) from their phylogenetic analyses. Omissions like this are why independent research using wide gamut analyses are necessary to clear out old cobwebs and new myths. Attempts at suppression of the LRT were cited by Müller at ResearchGate.
References Ezcurra MD et al. (17 co-authors) 2020. Enigmatic dinosaur precursors bridge the gap to the origin of Pterosauria. Nature https://doi.org/10.1038/s41586-020-3011-4 Müller RT 2022. The closest evolutionary relatives of pterosaurs: what the morphospace occupation of different skeletal regions tell us about lagerpetids. The Anatomical Record. https://doi.org/10.1002/ar.24904 Müller RT and Garcia MS 2022. Oldest dinosauromorph from South America and the early radiation of dinosaur precursors in Gondwana. Gondwana Research https://doi.org/10.1016/j.gr.2022.02.010 Novas FE and Agnolin FL 2016Lagerpeton chanarensis Romer (Archosauriformes): A derived proterochampsian from the middle Triassic of NW Argentina. Simposio. From Eventos del Mesozoico temprano en la evolución de los dinosaurios”. Programa VCLAPV. Conferencia plenaria: Hidrodinámica y modo de vida de los primeros vertebrados. Héctor Botella (Universitat de València, España) 2016
Publicity form Müller and Garcia 2022: The struggle was real for the oldest proto-dinosaur found in South America. Dinosaur ancestors weren’t so “dino.” By Elizabeth Rayne online here.
Turns out the oldest overlooked proto-dinosaur, PVL 4597 (Fig 1), is also from South America.
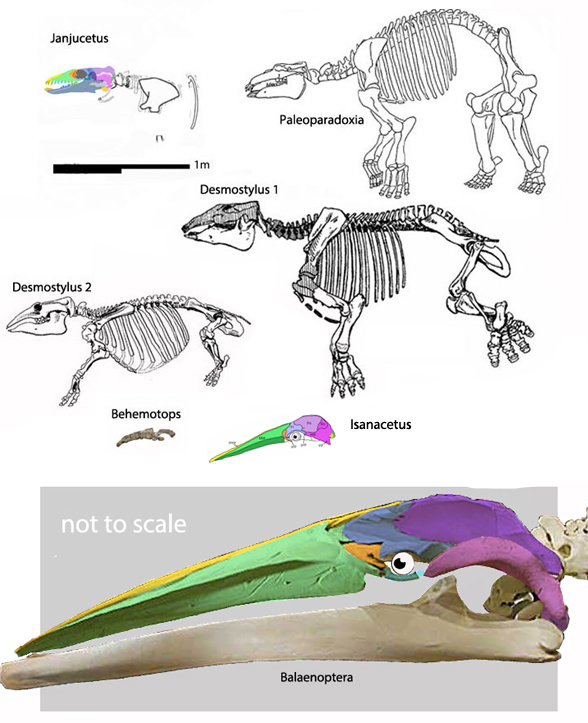
Few placental clades are as misunderstood as the Desmostylia. This was well documented by Domning, Ray and McKenna 1986 with their review of desmostylian origin (Figs 1, 3) and interrelation studies (abbreviated below). This timeline provides insight into the phylogenetic struggles paleontologists suffered over this clade in the pre-cladistic, pre-software era.
Figure 1. Isanacetus compared to scale with desmostylian sisters recovered in the LRT. Balaeonoptera (at bottom) is much reduced.
Unfortunately, this struggle continues in 2022 due to… (drum roll, please) …taxon exclusion. You’ll either weep or chuckle with bemusement as the best PhDs of their respective eras kept their blinders on, spending decades struggling for an answer that wasn’t going to be found where they were looking. Apparently most workers were hesitant to look somewhere other than where others had looked for a solution to the 134-year-old desmostylian problem. Note how professors through the decades kept beating around the same bush without success or resolution, not imagining they should be looking elsewhere.
Abridged from Domning, Ray and McKenna 1986: “O.C. Marsh (1888) referred Desmostylus to the Sirenia.”
“Yoshiwara and Iwasaki (1902), believed their find [a desmostylian] to be some sort of proboscidean, based in part on a letter from H.F. Osborn. Although Osborn had “informed” them that the skull belonged to a proboscidean, Yoshiwara and Iwasaki demonstrated that it was not like deinotheres or elephantids and therefore would have to represent a branch from the primitive proboscideans, near the origin of that order from among the other ungulates. They also mentioned some similarities to Sirenia.”
“Hay (1915), followed by Matsumoto (1918), placed the Desmostylidae in the Sirenia, although he emphasized that the Desmostylidae were very different from other (true) sirenians.”
“Abel (1922:381; 1923) abandoned his view of proboscidean affinities of Desmostylus in favor of a bizarre notion that it belonged to the mammalian subclass Allotheria (= Multituberculata).”
Although misguided and wrong, at least Abel was expanding his gamut.
“Sickenberg (1938), however, argued strongly against a desmostylian-sirenian relationship.”
Ijiri (1939) considered Desmostylus to be an ungulate “in the broadest sense” but not a monotreme, multituberculate, marsupial, or sirenian.”
Minkoff (1976) suggested that desmostylians should be placed with the Amblypoda rather than the Paenungulata.
Amblypoda are basalmost condylarths = Pantodontids and Phenacodontids in the LRT. Like others before, Minkoff was likewise playing darts in the dark by suggesting and guessing.
Domning, Ray and McKenna 1986 correctly compared desmostylians with hippos, but ironically did not permit hippos to enter analysis. The authors made no mention of the mysticeti, mesonychids, or oreodonts, which turn out to be related to desmostylians when more taxa are added to analysis, as in the large reptile tree (LRT, 2061 taxa, subset Fig 2).
Figure 2. Subset of the LRT focusing on condylarth placentals including desmostylians nesting basal to mysticetes, the one clade desmostylian workers universally and historically seem to avoid.
But wait, there’s more… Workers often and correctly associate desmostylians with anthracobunids, but once again, fall into the same proboscidean sticky trap.
Domning, Ray and McKenna 1986 considered Anthracobune a proboscidean, so their phylogenetic problems extended beyond desmostylians. In the LRT (Fig 2) anthracobunids nest between hippos and desmostylians, far from elephants.
From Domning, Ray and McKenna 1986: “Manning’s identification of Anthracobune as a Moeritherium-Vike animal was also generously made known to R.M. West, who was the first to publish on the matter (West, 1980:518; 1983). West placed Anthracobune in the Moeritheriidae.”
“During the long interval from 1940 to 1980, Anthracobune (with “Pilgrimella”) had masqueraded as an artiodactyl (Pilgrim, 1940; Gingerich, 1977; Coombs and Coombs, 1977; and most other authors), a perissodactyl (Coombs and Coombs, 1977:303; 1979), and a phenacodontid condylarth (Van Valen, 1978, fig. 3). Wells and Gingerich (1983) assigned it to a new family Anthracobunidae within the Proboscidea, and (based on an examination of the specimens of Behemotops reported herein) suggested that the Desmostylia, as well as the Moeritheriidae and the Sirenia, may be derived from anthracobunids. The fact that Anthracobune occurs in southern Asia, rather than in Africa, was doubtless a major cause of its long neglect in discussions of the phylogenetic origin of proboscideans and their possible affinities to the desmostylians.”
Lessson 1: Let your software and a wide gamut taxon list tell you which taxa to include, not the other way around. That would be considered akin to gerrymandering or cherry-picking. And it’s okay to make mistakes along the way. Look at the professionals of the past that kept their blinders on despite their lack of confidence in their results (see above). None were ostracized or lost their paycheck or place in science.
Lesson 2: Don’t depend on dental traits at first, not until your wide gamut cladogram is fully resolved and major clade interrelationships are confidently known from substantial skulls and skeletons. Then you can drop in your partial taxa and dental taxa with greater confidence.
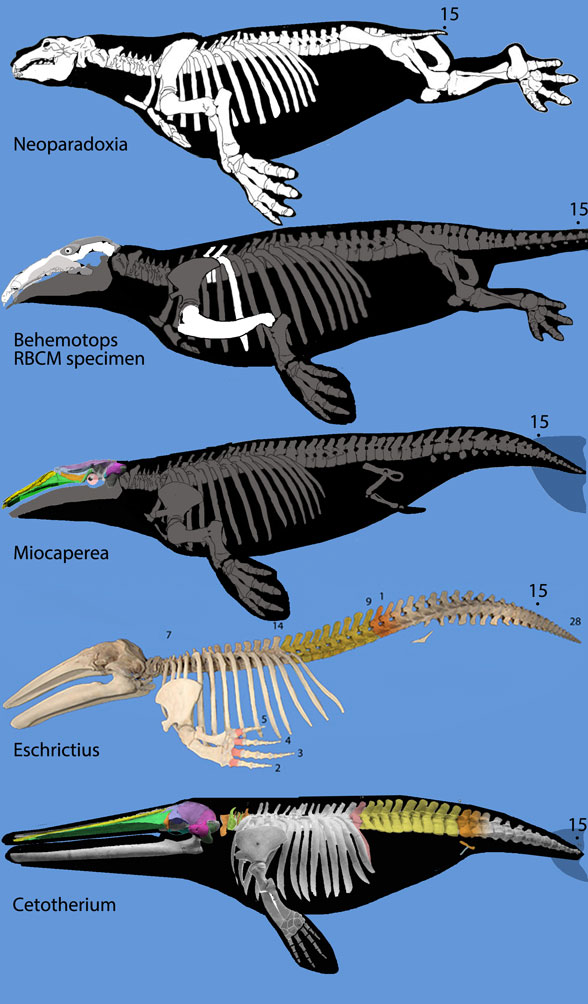
Figure 3. Rorqual evolution from desmostylians, Neoparadoxia, the RBCM specimen of Behemotops, Miocaperea, Eschrichtius and Cetotherium, not to scale.
If you are at all prescient, or a long time reader, you’ll know what comes next: Pterosaurs. Turtles. Snakes. Reptiles. Placoderms. Creodonts. Bats. Catfish. Diapsids. These are all taxa that find a good, secure (= fully resolved) home in the LRT, something they cannot presently find in university textbooks. Desmoystlians are also firmly nested in the LRT, but still an enigma otherwise.
Remember, all professors were once students eager to please their own professors by parroting paradigms. All academic authors know they have to satisfy anonymous referees (= sometimes jealous and self-serving academic competitors). Both systems discourage discovery and heresy. In academia consensus rules (see timeline above). It always has. And it always will. That means it comes down to politics, not science. That’s why it sometimes takes an outsider to pull the curtain back on enigma taxa to give them a home based on the scientific method with complete transparency and falsifiability …and to take the occasional barb for the trouble of doing so without payment or tangible reward.
References Domning DP, Ray CE and McKenna MC 1986. Two New Oligocene Desmostylians and a Discussion of Tethytherian Systematics. Smithsonian Contributions to Paleobiology 59:56pp. Peters D unpublished. The triple origin of whales. PDF on ResearchGate.org
Phosphatherium escuillei(Gheerbrant, Sudre and Cappetta 1996; Late Paleocene; 10cm skull length; Fig 1) has been a traditional elephant ancestor since its discovery (Gheerbrant, Sudre and Cappetta 1996) and rediscovery as a second specimen (Gheerbrant E et al. 2005; Gheerbrant 2009) assigned to the same genus and species.
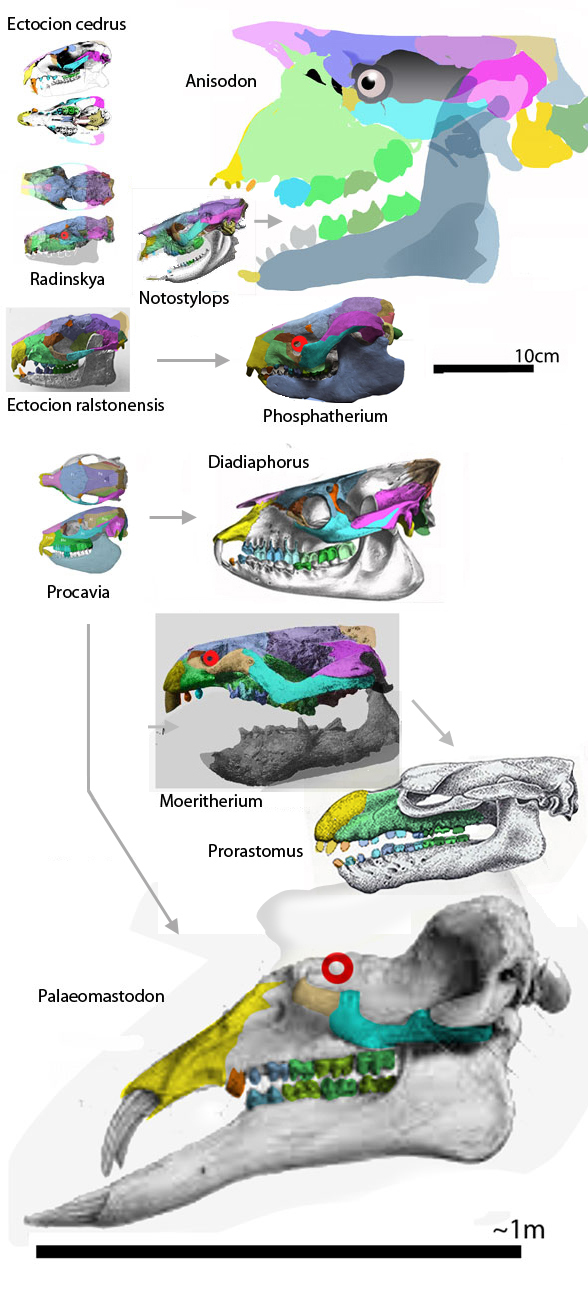
After testing in the large reptile tree (LRT, 2062 taxa, subset Fig 3) both new Phosphatherium specimens nest together, but derived from the more plesiomorphic, Ectocion ralstonensis (Fig 1), a taxon omitted by prior authors. That means Phosphaterium is less of an elephant ancestor than is the omitted, more plesiomorphic Late Paleocene taxon, Ectocion ralstonensis (Cope 1882 a, e; Granger 1915; Thewissen 1990). “A Palaeocene proboscidean from Morocco” got published in Nature in 2005. I’m guessing, “An Ectocion descendant leaving no descendants” would not have gotten published in Nature. Ya gotta do, whatcha gotta do in paleo.
In the LRT taxa closer to elephants include Procavia, the hyrax, basal to sirenians. These two taxa and their descendants have a single pair of incisor tusks at most.
Figure 1. Above: Phosphatheirum. Middle: Ectocion. Below: Eritherium parts applied to a ghosted image of Ectocion.
Here two Phosphatherium skulls (Fig 1) are not the same species, but share more traits with each other than with other taxa.
Figure 2. Fossils in the elephant + manatee clade including Phosphatherium Palaeomastodon, Moeritherium, Ectocion, Notostylops, Radinskya, Diadiaphorus, Procavia and the chalicothere, Anisodon.
Phosphatherium was close to the ancestry of elephants (Fig 2) but several taxa were closer, despite the big bone appearance of Phosphatherium, convergent with elephants.
Figure 3. Subset of the LRT focusing on terrestrial placentals including Phosphatherium nesting in a clade between hyraxes and manatees + elephants.
Gheerbrant 2009 described the partial remains of Eritheriumazzouzorum, (Fig 1) as “the oldest and most primitive elephant relative” at 60mya, 5 million years earlier than Phosphatherium. The partial remains consist of a palate (maxilla + jugals) and teeth, plus paired frontals and nasals. This is too little to fragmentary for the LRT, but note how well the pieces fit into a coeval Ectocion ralstonensis bauplan (Fig 1). Here the subhead, “Also a hyrax and manatee relative” would have been appropriate. The LRT lists elephant ancestors back to the Cambrian, just like every other included taxon.
Gheerbrant 2005wrote: “We report here significant new material belonging to the oldest and most primitive known Proboscidean, Phosphatherium escuilliei Gheerbrant, Sudre & Cappetta, 1996, from the early Eocene of the Ouled Abdoun phosphatic basin, Morocco. This material permits the first reconstruction of the skull and most of upper and lower dentition of Phosphatherium escuilliei.”
This second specimen (Fig. 1) does bring much needed data to this genus.
“The species, which is one of the oldest and most primitive known representatives of modern orders of ungulates, becomes one of the best known among them. Its dentition shows a noticeable dental variability, which is interpreted, at least provisionally, to be intraspecific.
Elephants don’t have diverse post-incisor tooth morphology like more primitive taxa do.
“The skull of Phosphatherium escuilliei is very primitive in many respects. It is long with an elongated facial part and a narrow rostrum. The toothrow does not extend posteriorly beyond the middle of the skull. The nasals are long and located anteriorly (i.e. nasal fossa not retracted). There is no contact between the premaxilla and frontal. There is a strong postorbital constriction and a distinct postorbital process on the frontal. The zygomatic arches are noticeably expanded laterally.
That lateral expansion is not shared with Ectocion, sirenians or elephants.
“The sagittal and nuchal crests are strong. The external auditory meatus is open ventrally. The braincase is strongly compressed laterally”.
In other words, the brain was not wide or large.
“Some primitive features of the dentition are also noticeable.“
That means distinct incisors, canines, premolars and molars.
“However, Phosphatherium escuilliei displays several strikingly advanced features, especially proboscidean and tethytherian features. A cladistic study of 129 features of Phosphatherium escuilliei supports the monophyly of Proboscidea and the inclusion of Phosphatherium within the order.
The LRT does not confirm the inclusion of Phosphatherium within the clade of elephants.
“The most significant Proboscidean synapomorphies found in Phosphatherium are: 1) the well developed zygomatic process of the maxillary which contributes significantly to the ventral border of the orbit and to the zygomatic arch; 2) the relatively large size of the pars mastoidea of the periotic; and 3) the hypoconulid in a labial position (a state unique to Proboscidea).“
The Gheerbrant et al. 2005 taxon list included Ectocion (but which species?) representing the suprageneric taxon ‘Phenacodontidae’. The authors mistakenly included the unrelated suprageneric clades, Embrithopoda, Anthracobunidae and Desmostylia, erroneously following current textbooks and traditions. The authors employ Perissodactylia, a related clade, but again it is used as a suprageneric taxon. That’s always dangerous and rife with bias and cherry-picking. The authors employ a long list of dental traits not used by the LRT.
By contrast the LRT employs no suprageneric taxa and no chimaera taxa, only specimens and species. That’s why both Phosphatherium taxa (Fig. 1) entered the LRT separately. In the LRT there is no cherry-picking of data from suprageneric taxa to suit an author preference.
The omission ofEctocion ralstonensis from prior analyses needs to be repaired in future studies on elephant origins.
The Phosphatherium authors employed taxa not tested in the LRT. One: Late Paleocene Phenacolophus is a mandible + teeth taxon.
Two: Late Paleocene Minchenella (= Conolophus, preoccupied by an iguana) was described by Domning, Ray and McKenna 1986 as “a suitable candidate to be the common ancestor of both the Desmostylia and the Proboscidea” based on a molar cusp. The LRT separates these two clades based on more complete skeletons. More on that issue tomorrow.
References Domning DP, Ray CE and McKenna MC 1986. Two New Oligocene Desmostylians and a Discussion of Tethytherian Systematics. Smithsonian Contributions to Paleobiology 59:56pp. Gheerbrant E, Sudre J and Cappetta H 1996. A Palaeocene proboscidean from Morocco. Nature. 383 (6595): 68–71. Gheerbrant E et al. 2005. Nouvelles données sur Phosphatherium escuilliei (Mammalia, Proboscidea) de l’Éocène inférieur du Maroc, apports à la phylogénie des Proboscidea et des ongulés lophodontes. Geodiversitas 27 (2), 2005, pp. 239-333. Gheerbrant E 2009. Paleocene emergence of elephant relatives and the rapid radiation of African ungulates. Proc Natl Acad Sci USA. 106(26): 10717–10721.
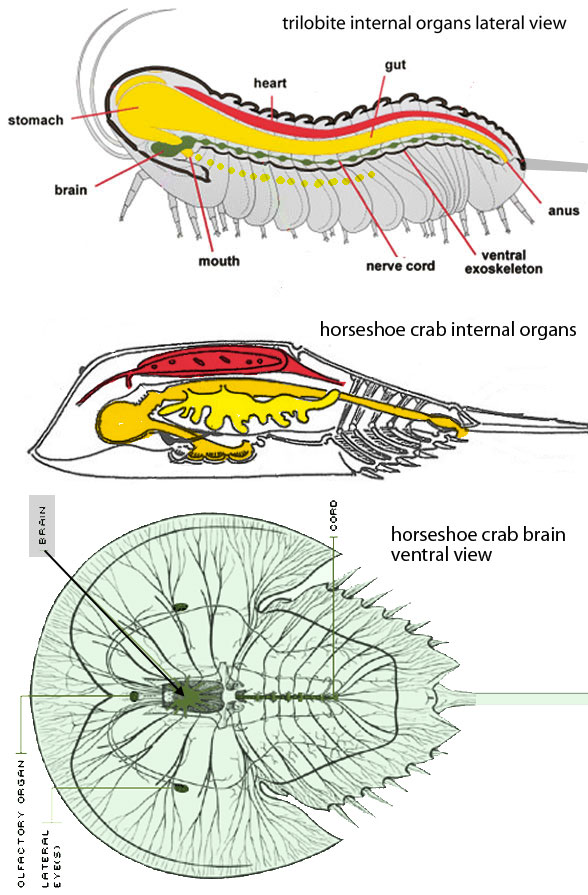
Here is where I go ahead and state the unaccepted obvious: Horseshoe crabs (Figs 1–4) are living trilobites (Figs 1–4). Everyone already senses this at some basic level. This is especially so for that clade of Early Cambrian trilobites sporting a long, sharp telson (= axial spine) rather than a more traditional wide pygidium. That telson-sporting clade is represented here by Pseudosaukianda (Figs 1, 3).
The University of Houston describes how most universities think about horseshoe crabs and trilobites: “The nearest thing to a trilobite today is the horseshoe crab with a very similar exoskeleton. Trilobites lasted over 300-million years and finally died out not long before dinosaurs arose.”
U of Houston represents the consensus: that trilobites died out.
Wikipedia repeats that adage when they report,“Trilobites have no known direct descendants. Though horseshoe crabs are often cited as their closest living relatives, they are no closer evolutionarily than other chelicerates.”
There was a time not so long ago when teachers used to tell their students dinosaurs are extinct. Then teachers had to correct themselves in the 1990s when monophyletic clades based on software-assisted phylogenetic analysies suddenly became ‘a thing’. And China began producing feathered dinosaurs and toothy birds by the hundreds. Sadly, all that ‘new insight’ arrived over a hundred years after Archaeopteryx was known to early adopters as a transitional dinosaur-to-bird.
Here online photos and graphics are collected that you can show to your trilobite-loving friends… after first rousing yourself from your own paradigm slumber to digest and accept this tiny step away from tradition and consensus.
Figure 1. Early Cambrian Pseudosaukianda and Limulus. the extant horseshoe crab. In 540 million years these two have changed far less than our coeval chordate ancestors.
Trilobites evolved a variety of shapes and sizes. Among them is the Early Cambrian genus, Pseudosaukianda (Fig 1), one with a telson. That trilobite genus generally matches the overall morphology of Limulus, the extant horseshoe crab (Figs 1, 3) and its transitional ancestors (Fig 2). Turns out that trillobite telson is retained in Limulus. It’s not there by convergence. There are no other taxa that have such a telson. None are closer to Limulus than Pseudosaukianda and its transitional kin (Fig 2).
But wait. There’s more. Much more.
Figure 2. Horseshoe crab ancestors demonstrating the fusion of thorax elements. The earliest horseshoe crab traditionally post-dates Pseudosaukianda by 100 million years. It’s time for a paradigm shift.
In dorsal view (Fig 3) both Pseudosaukianda and Limulus have similar morphologies with a similar number of lateral spines arising from the thorax, whether fused or not. In Limulus every other thorax segment evolves into a narrower lateral spine between wider lateral spines. Not much change here considering the half a billion years that separates these two.
Figure 3. In dorsal view Pseudosaukianda and Limulus share the same bauplan. In the horseshoe crab the thorax elements are now fused. That’s all and that’s not much for 540 million years of evolution.
In ventral view both Pseudosaukianda and Limulus also have similar morphologies. Horseshoe crabs lose the gill element from the first six limbs and lose the limb element from the remaining gill plates. Both taxa have a ventral and central oral cavity between the bases of six sets of legs that all contribute to grabbing and processing food items before swallowing. This is not the way taxa derived from segmented annelids with a terminal mouth acquire food. It is the way primitive marine flatworms with a single central mouth/anus opening acquire food. Give a flatworm legs, armor and an anus and pretty soon you can call it a trilobite, as reported earlier here.
Figure 3. Trilobite limbs are rarely preserved, so Triarthrus limbs are modified here to fit the shorter morphology of Pseudosaukianda. The loss of limbs or gill plates is really the only difference between Pseudosaukianda and Limulus. Note how trilobites feed like flatworms. They cover their food items and use their proximal legs to draw it forward into the posteriorly-oriented ventral oral cavity. In horse shoe crabs this oral cavity is even more central to the leg bases.
Antennae Horseshoe crabs lack antennae (Fig 3). Very few trilobites preserve antennae, legs + gills. So the reduction and loss of antennae between trilobites and horseshoe crabs has not been documented due to taphonomic (= preservation) issues.
In parasagittal view (Fig 4) both the horseshoe crab and trilobite have a stomach filling or entering the anterior of their cephalon. The ventral mouth is posterior to this organ and is oriented posteriorly. A simple intestine extends to the posteroventral anus, opening beneath the base of the telson. The elongate heart/circulatory system is dorsal to the intestine. The brain + nervous system is ventral. It’s a pretty good match, both inside and outside. No other taxa are a closer match.
Figure 4. Triolbite and horseshoe crab in lateral view showing stomach (crop) in front of the brain, mouth opening posterior to the stomach, small brain ventral to these elements. All this is more like a marine flatworm than an arthropod. Crustaceans and insects are descendants of trilobites. Long narrow velvet worms and shorter tardigrades arise from nematodes (= round worms) and annelidids (segmented round worms).
If any other similar animal taxon has any part of its stomach in its head, let me know. Likewise, if any other animals acquire and masticate food between the toothy ventral bases of their legs in the middle of their body, let me know. I am aware that crustacean, arachnid and insect mouth parts are former legs, but the feeding structures are all under a much smaller head in those taxa. When and if eventually tested here they will be compared.
It’s all about monophyly. Horseshoe crabs are living trilobites,just like birds are theropod dinosaurs and humans are primates, placentals, mammals, amniotes, tetrapods, vertebrates and chordates. If you understand the concept of monophyletic clades, you’ll understand this apparently novel and long overdue hypothesis of interrelationships in trilobites. In evolution every living taxon is somehow connected to every other taxon. Separating for teaching purposes is appropriate. So is lumping.
This insight was low-hanging fruit that should have been plucked decades ago. The practice of counting legs in arthropods (e.g. insects have six, spiders have eight) may be to blame here for the academic separation of Limulus from Pseudosaukianda, when they should have been lumped together as they are here. The opposite problem exists in diapsid-grade taxa that are not related to one another.
It is also important to not use suprageneric taxa. Most trilobites are not appropriate outgroups for horseshoe crabs.
Please forgive this venture beyond chordates. It was an issue that needed to be addressed.
The following horseshoe crab video sheds more light on this newly minted trilobite. You’ll see the mouth central to the limbs in this inverted specimen. You’ll see the legs are now distinct from the gill plates. You’ll see this horseshoe crab curl up like a trilobite. You’ll see how harmless it is. You’ll see the primitive eyes and sex organs all presented by a knowledgeable handler. See video here. No preview was permitted. https://www.youtube.com/watch?v=rmlOAlodt54
One thing you won’t see is how horseshoe crabs swim, which is rarely. They do the backstroke. Here’s a video on that:
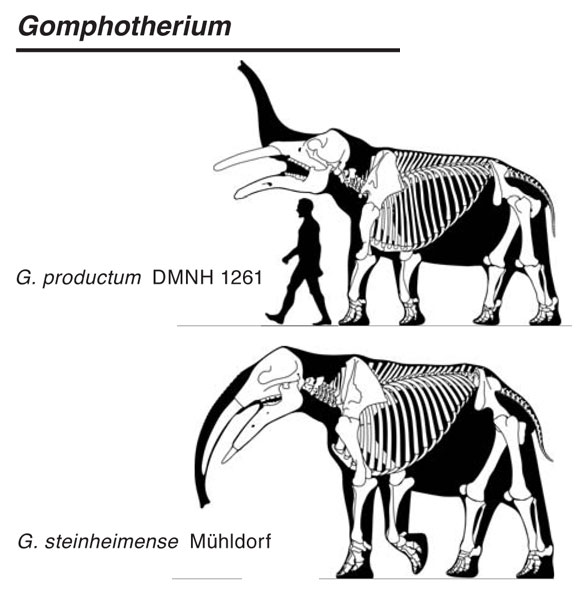
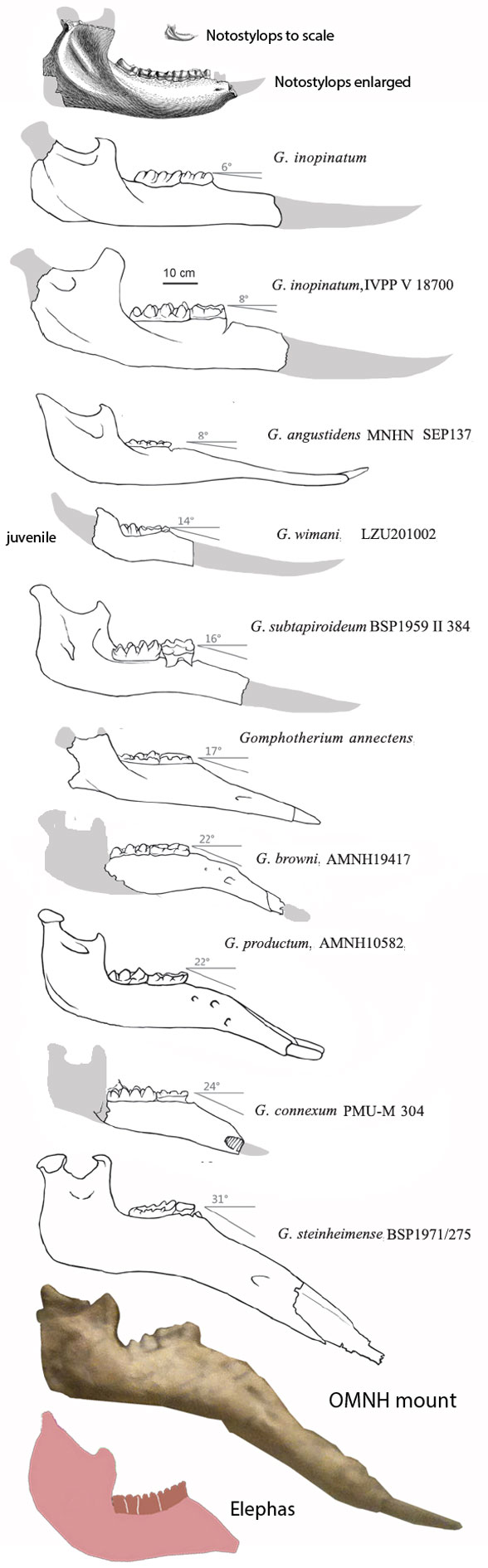
Known since Kaup 1829, Deinotherium giganteum(Kaup 1829; Late Miocene; Figs 1, 2) is a large deinothere (Fig 2), the clade of elephants lacking premaxillary tusks. Instead large dentary (= mandible) tusks curl ventrally then posteriorly in all deinotheres (Fig 2) , distinct from all other elephant genera, as everyone already knows. Interrelationships have been traditionally murky since transitional taxa have not yet been documented.
Figure 1. Skull of Deinotherium in two views. Colors added here. Note the enormous naris visible in dorsal view. The nasals are either absent or fused to the frontals in this diagram.
Figure 2. Deinotherium figures from Larramendi 2016.
Larramendi 2016 published several excellent lateral view skeletons of deinotheres (Fig 2) and gomphotheres (Fig 3) along with several other elephant clades.
Larramendi 2016calculated body mass in extinct taxa “by the by the Graphic Double Integration volumetric method which is based on technical restorations from graphical reconstructions of fossils employing photos, measurements and comparative anatomy of extant forms. The method has been tested on extant elephants with highly accurate results. From the shoulder heights, several equations were created to find out the body mass of a series of extant and extinct species. A few of the largest proboscideans, namely Mammut borsoni and Palaeoloxodon namadicus, were found out to have reached and surpassed the body size of the largest indricotheres.”
In the LRT (Fig 5) indricotheres are giant three-toed horses. Horses and rhinos are hyracodontids. Those are tapir relatives. Those are chalicothere and hyrax relatives. Those are Ectocion relatives. Those are artiodactyl relatives. Those are oreodont and mesonychid relatives. Etc.
Figure 3. Gomphotherium, like Deinotherium, had a longer dentary and dentary tusks.
A recent paper by Baleka et al. 2022 produced a phylogenetic analysis of elephants using total evidence (genes + traits). They wrote: “The order originated around 60 million years (Ma) ago in Africa. At present, the oldest proboscidean fossil is Phosphatherium escuilliei Gheerbrant, Sudre & Cappetta,1996, from Morocco. It comprises cranial and mandibular elements dating to 55 Ma (Gheerbrant, 2009).
We will take a closer look at Phosphatheriumfrom Paleocene Morocco in the near future. Turns out it’s not a direct elephant ancestor (Fig 5) in the LRT.
Baleka et al. 2022 continue: “The evolutionary history of proboscideans is marked by three major radiations.”
“The first occurred during the late Palaeocene/Eocene, with the diversification of primitive proboscideans.”
“The second radiation took place during the early Miocene, with the diversification of ‘‘Gomphotheriidae’’ Hay, 1922, (used sensu lato throughout the text, indicated by brackets) Mammutidae Hay, 1922, and Stegodontidae Osborn, 1918, a family within the Elephantoidea.”
“The last radiation took place during the late Miocene/early Pliocene and resulted in the diversification of Elephantidae Gray (1821), as well part of the superfamily Elephantoidea, including the living elephants.”
The LRT indicates elephant ancestors do not include Phosphatherium, but arise directly from Procavia-llike and Diadiaphorus-like tusked taxa.
Figure 4. Gomphotherium jaws. Note the elongate dentary and large dentary tusks. Those are not seen in other elephant clades.
Baleka et al. reported, “The historical biogeography analysis (Figures 4, S4, and S5) suggests that proboscideans may have left Africa only three times in phylogenetically distant clades (Deinotherium or Deinotheriidae, Mammutidae, and Elephantida).”
The LRT nests Elephas with Mammuthus and Deinotherium with Gomphotherium. Mammut (the mastodon) has not yet been tested.
Figure 5. Subset of the LRT focusing on elephants and their relatives. As in Baleka et al. 2022, Deinotheres are close to Palaeomastodon. Distinct from Bakeka et al. Gomphotherium nests with deinotheres here, rather than closer to extant elephants than Mammut the mammoth. Be wary of DNA studies.
References Baleka et al. 2022. Revisiting proboscidean phylogeny and evolution through total evidence and palaeogenetic analyses including Notiomastodon ancient DNA. iScience 25, 103559 https://doi.org/10.1016/j.isci.2021.103559 Kaup JJ 1829. Neues Säugethier, Deinotherium: Deinotherium giganteum. Isis 22(4):401–404 Larramendi A 2016. Shoulder height, body mass, and shape of proboscideans. Acta Palaeontologica Polonica 61 (3):537–574.
Vanellus miles novaehollandiae (Boddaert 1783; up to 37cm in length; Figs 1, 2) is the extant masked lapwing, aka spur-wing plover, of Australia and New Zealand, notable for transforming manus digit 1 into a formidable spike, and for those yellow facial wattles. Otherwise it is almost identical to the more plesiomorphic Charadrius. the plover, with which it nests in the LRT.
Figure 1. Vanellus the extant spur wing is not related to other spur-wings in the bird clade.
Figure 2. Skeleton of Vanellus with insets for the left and right manus. Digit 1 is colored red here.
Comparisons to the horned screamer, Anhima (Fig 3), are warranted because both bear manus spurs. Analysis indicates the two do not nest together and the morphology of the spur arises from different bones.
Figure 3. Horned screamer (genus: Anhima) skeleton. Spiked manus is is purple. Note, the spike is separate and distinct from digit 1 here. Compare to figure 2.
Convergence is demonstrated in this example of two unrelated birds with hand spurs.
References Boddaert P 1763.The Histoire Naturelle, générale et particulière, avec la description du Cabinet du Roi (Natural History, General and Particular, with a Description of the King’s Cabinet)