If you follow published drawings of the armored Ordovician lancelet, Arandaspis (Fig 1), you’ll find phylogenetic frustration. No other related taxa have eyeballs oriented anteriorly like Jeep headlights.
Figure 1. Arandaspis diagram. Colors and second frame with face lift added here.
However, if you flip up the face, rotating the eyeballs dorsally, then you’ll find a long list of homologies with otherwise similar (and therefore related) taxa, like Metaspriggina (Fig 1) a shorter, deeper, descendant of lancelets and lampreys. Here (Fig 1) both up and front face options are presented. Lampreys and Metaspriggina have a single median naris between the eyeballs. With the proposed orientation of the eyeballs, so would Arandaspis.
Metaspriggina walcotti (Simonetta and Insom 1993; Morris and Caron 2014; Cambrian, 500 mya; up to 10cm) is an early chordate, naked, jawless and finless, but with two anterodorsal eyes and a naris between them. The orbits signal the genesis of a proto-skull. The atrium was smaller and the swimming muscles were larger than in lancelets, so this was probably a mobile feeder, still without fins, using its eyes to seek prey and avoid predators. Gill slits opened directly, as in lampreys, without an atrium. Descendants, like Arandaspis (below), were basal to armored placoderms. Our ancestors were the ones without such armor according to the large reptile tree (LRT, 2130 taxa)
Figure 2. A partial and still buried fossil of Arandaspis. Too little is presented here to be sure of orientation, but it appears to be presenting its left side here. What are those green dorsally-oriented semi-circles at left?
Arandaspis prionotolepis (Ritchie and Gilbert-Tomlinson 1977; Ordovician, 465mya; ~15 cm long) is a member of the Arandaspididae. It looked like a large, armored tadpole, or a large armored Metaspriggina with a tail fin. This jawless, filter feeder was basically an armored lancelet. Both eyes and nostrils probably faced upward (based on evidence (Fig 2) and phylogenetic bracketing, see above) cantilevered above and hanging over the jawless oral opening, as in hagfish. Several armored gill slits appeared between the dorsal ‘carapace’ and ventral ‘plastron’.
References Ritchie A and Gilbert-Tomlinson J 1977. First Ordovician vertebrates from the Southern Hemisphere. Alcheringa 1:351-368. Simonetta AM and Insom E 1993. New animals from the Burgess Shale (Middle Cambrian)and the possible significance for the understanding of the Bilateria. Bolletino Di Zoologia 60:97–107.
Take a look at this new video from PBS Eons and you’ll understand my replies below:
Figure 1. Click to play PBS Eons video, but be ready for outdated information
Reply 1: Mystacodon (Fig 1) is not a mysticete nor a mysticete ancestor. The list of mysticete ancestors includes desmostylians, anthracobunids, hippos, mesonychids and oreodonts (Fig 2). Odontocete ancestors do include Indohyus, as shown in the video, and also include echo-locatiing tenrecs still living in Madagascar, which was once connected to Pakistan, home of Pakicetus, the walking whale and giant tenrec.
Figure 1. This toothy whale with a tiny pelvis is Mystcodon, originally promoted as the earliest known mysticete (baleen whale).
Reply 2: Maiabalaena is indeed a toothless mysticete, one of the cetiotheres, known for their small size and oversize head. Be careful with traits like this. A taxon could be in the process of evolving baleen, or in this case, losing baleen its ancestors once had.
Figure 2. Origin of the Mysticeti according to the LRT.
Janvier 2008 reported on early jawless vertebrates, but did not understand that the dorsal narial opening of lampreys is homologous with the dual pair of nares seen in sturgeons (Fig 4). Janvier reported, “The dorsal, median, asohypophysial complex of osteostracans has been regarded as identical and homologous to that of lampreys, but recent investigation (notably on the galeaspid braincase) now suggests that this resemblance is in fact a convergence.”
In the LRT (Fig 3) galeaspids are basal to the placoderm clade not to the sturgeon clade. Lampreys (= Pteromyzon) are basal to both.
Figure 1 Pharyngolepis oblongus from Janvier 2008. Colors added here.
By contrast, The large reptile tree (LRT, 2130 taxa, subset Fig 3) indicates the dorsal naris in lampreys is indeed homologous with the same opening in osteostracans. Janvier’s images of Pharyngolepis oblongus (Fig 1) show a single dorsal narial opening expanding laterally. Subsequently more lateral movement splitting the nares in two, followed by additional splitting splitting the two into four (incurrent and excurrent, left and right) continues in tiny Early Cambrian Haikouicthys (Fig 5) and concludes in the sturgeon, Acipenser (Fig 4). A valid wide-gamut cladogram resolves all phylogenetic problems.
Figure 2. Pharyngolepis relatives leading to gnathostomes to scale and full scale on a 72dpi monitor. Note the tiny size of Haikouichthys. Either this is phylogenetic miniaturization or all fish were that tiny in the Early Cambrian.
Before these basal vertebrates evolved jaws and pelvic finss (Fig 3), they developed pectoral fins by convergence with placoderms and pre-gnathostomes. In the Pharyngolepis clade a large anal fin developed not found on the other lineage leading to gnathostomes until Thelodus (Fig 2) and sturgeons came along.
Figure 3. Taxa in the Pharyngolepis clade to scale and in phylogenetic order.
The addition of Pharyngolepis helped refine prior scorings in the LRT. Because this taxon nested at the base of its clade, that means it was very close to the lineage of gnathostomes, bony fish, tetrapods and ultimately humans.
Figure 3. Subset of the LRT focusing on basal vertebrates including Pharyngolepis derived from lampreys and Metaspriggina. Note: Early Cambrian Haikouichthys nests basal to sturgeons, an earlier appearance of fish of this grade than recovered previously.
I relied on the drawings and diagrams of prior workers to make the MacClade scores. Coloring the bones in their tetrapod homologs greatly helped understand which dermal splinters would ultimately fuse and which would someday split, as in all other vertebrates.
Figure 4. How one wide nostril on Pharyngolepis splits into four in the juvenile sturgeon, Acipenser.
Pharyngolepis documents how skull bones first appeared in pre-gnathostomes. Earllier,Birkenia held that honor. Placoderms did their own thing. So did galeaspids. Note the absence of the marginal tooth-bearing bones plus their hinge, the quadrate. Those appear later, in nurse sharks. The great size of the nasal complex was probably related to prey acquisition via olfaction, as in later sharks and earlier lampreys.
Figure 5. Haikouichthys in situ and skull closeup, colors added. Lower image shown 5x life size. See figure 6 for an overall view and comparisons.
What about the transitional taxon from the Early Cambrian, Haikouichthys? It’s hard to tell what’s going on here (Fig 5), but take a look at Haikouichthys in figure 2. At 2.5cm, it could be yet another phylogenetically miniaturized transitional taxon. Either that or all Early Cambrian fish were that tiny. Tiny is where new structures have their genesis, in this case the splitting of a single naris into four. Haikouichthys is not strongly ossified. It doesn’t need to be and it cannot be because it is so tiny. That’s where a single median naris likely split into four nares (two incurrent immediately followed by two excurrent, left and right) for better direction-finding of trace odors in sea water. Little Silurian Thelodus is a transitional taxon between Haikouichthys and sturgeons, but preserved ventrally exposed, so the dorsal nares are hard to see.
Figure 6. Haikouichthys in situ and compared to a juvenile sturgeon. Colors added. White arrow points to oral entry. Orange arrow points away from anal exit between pelvic fins. Shown 3.6x life size. See figure 5 for a close-up of the skull.
The Early Cambrian appearance of tiny Haikouichthys means precursor taxa and phylogenetic splits in the LRT (Fig 3) preceded the time of Haikouichthys in the Early Cambrian. That also means the genesis of sturgeon-grade fish likely extends back to the Middle Cambrian, a novel hypothesis that needs confirmation with fossil evidence of sturgeons, or at least thelodonts, in the Middle to Late Cambrian.
Haikouichthys ercaicunensis (Luo, Hu & Shu 1997; Shu et al.1999; HZf-12-127; 2.5cm) is an Early Cambrian transitional taxa basal to sturgeons in the LRT. The pectoral fin is not separate from the body. A prominent dorsal fin is present. So is a heterocercal tail. The series of posterior green diamonds are either Thelodus-like and sturgeon-like armor plates. Or those are ovaries and this tiny jawless fish is completely unarmored, like a sturgeon hatchling (Fig 6).
References Janvier P 2008. Early jawless vertebrates and cyclostome origins. Zoological Science 25: 1045–1056. Kiaer JA 1911. Skr. Vidensk. Christiania, Mat.-Naturv. Kl., 1911(7):1–19. Kiaer JA 1911. Skr. Vidensk. Christiania, Mat.-Naturv. Kl., 1911(7):1–19. Luo H. et al. 1997. New occurrence of the early Cambrian Chengjiang fauna from Haikou, Kunming, Yunnan province. Acta. Geol. Sin. 71, 97-104. Shu D-G et al. (8 co-authors) 1999. Lower Cambrian vertebrates from China. Nature 402:42-46.
Like Superman and Clark Kent, the phenacodont, Phenacodus, and the oreodont, Merycoidodon (Figs 1, 2), are never mentioned or seen together. The large reptile tree (LRT, 2129 taxa, subset Fig 3) sheds new light on their heretofor unknown interrelationship. In the LRT these two nest together.. and alone. Not sure why this was not recognized previously. It’s pretty obvious from all angles.
Figure 1. Merycoidodon compared to Phenacodus.
Yes, these two look different. But the LRT nests them together given the present taxon list of 2129. Phenacodus is the older, smaller and more primitive of the two.
Figure 2. Merycoidodon compared to Phenacodus.
Wikipedia reports, “Phenacodus is one of the earliest and most primitive of the ungulates, typifying the family Phenacodontidae and the order Perissodactyla.”Phenacodus had five digits on all four limbs. Merycoidodon and the oreodonts are not mentioned.
Wikipedia reports,“Merycoidodon is an extinct genus of herbivorous artiodactyl of the family Merycoidodontidae, more popularly known by the name Oreodon. The fore limbs had five toes (although the first one was vestigial), while the hind limbs had four.” Phenacodus is not mentioned.
Figure 3. Subset of the LRT focusing on herbivorous placentals. Agrichoerus and Promerychoerus are traditional oreodonts, but in the LRT are basal to their own clade of artiodactyls, just as the other oreodont, Merycoidodon, is basal to hippos and baleen whales.
Phenacodus primaevus (Cope 1873, Late Paleocene to Middle Eocene, 55 mya, 1.5m long) is one of the earliest and most primitive ungulates. Likely an omnivore, all four feet had five toes. All twenty claws were modified to become hooves. The middle finger and middle toe were the longest. Note the deep narial opening. No bones are fused. 44 teeth were present.
Merycoidodon culbertsoni (Leidy 1848; Late Eocene to Late Oligocene, 38–16 mya; 1.4 m in length)has been a traditional enigma, but nests here with Phenacodus at the base of the Mesonyx to mysticete (baleen whale) clade. Those two also nest at the base of the rest of the oreodonts at the base of the Homalodotherium clade + artiodactly clades. It is noteworthy that hippos do not nest with artiodactyls in the LRT, breaking a traditional paradigm. Merycoidon was a sheep-sized, short-legged herbivore living in herds that preferred well-watered areas. The forelimbs had five toes. The hindlimbs had four. The molars of these oreodonts were adapted for grinding, but the canines remained long as in mesonychids and hippos.
This appears to be a novel hypothesis of interrelationships. If not, please provide a citation so I can promote it here.
References Carrillo JD and Asher RJ 2017. An exceptionally well-preserved skeleton of Thomashuxleya externa (Mammalia, Notoungulata), from the Eocene of Patagonia, Argentina. Palaeontologia Electronica. https://palaeoelectronica.org. Cope ED 1873. Third notice of the extinct Vertebrata from the Tertiary of the Plains. Paleontology Bulletin 16:1-8. Cope ED 1884. Synopsis of the species of Oreodontidae. Paleontological Bulletin 38:503-572. Simpson GG 1967. The beginning of the age of mammals in South America. Part II. Bulletin of wthe American Museum of Natural History 137, 1-260. Leidy J 1873. Contribution to the extinct vertebrate fauna of the Western Territories. Geological Survey of the Territories 1. Leidy J 1848. On a new fossil genus and species of ruminantoid Pachydermata: Merycoidodon culbertsonii. Proceedings of the Academy of Natural Sciences of Philadelphia 4: 47–51. Leidy J 1850. [Abstract of remarks made before a meeting of the Academy of Natural Sciences of Philadelphia, December 17th, 1850]. Proceedings of the Academy of Natural Sciences of Philadelphia 5(1):121-122. Leidy J 1858. Notice of remains of extinct vertebrata, from the Valley of the Niobrara river collected during the exploring expedition of 1857, in Nebraska, under the command of Lieut. G K Warren, U. S. Top. Eng., by Dr. F.V. Hayden, Geologist to the expedition. Proceedings of the Academy of Natural Sciences of Philadelphia 1857: 20-29. Osborn HF 1898. Remounted skeleton of Phenacodus primaevus; Comparison with Euprotogonia. Bulletin of the AMNH10(9):159-164. Simpson GG 1980. Splendid Isolation, the Curious History of South American Mammals. Yale University Press, New Haven, CT. Thorpe MR 1921. Two new forms of Agriochoerus. American Journal of Science (8): 111–126.
Eleven lepidosaur experts worldwide came up short, by their own admission From the ‘In brief” paragraph: “Bolet et al. describe a second, more complete, specimen of the mid-Cretaceous genus Oculudentavis (new species) from the amber deposits of Myanmar. Comparative morphology and phylogenetic analyses prove that, despite its bird-like appearance, Oculudentavis is a bizarre lizard of uncertain affinities.”
‘Uncertain affinities’? That’s impossible. Everything evolves from something else.
Figure 1. Images of purported Oculudentavis from Bolet et al. 2020. Some colors added where necessary.
Don’t give up on your attempts at nesting taxa. Bolet et al gave up.Keep adding taxa to your own LRT until phylogenetic patterns emerge. Then add more taxa to refine your hypothesis of interrelationships. Everything evolves from something else.
Either that, or Google ‘Oculudentavis‘ to find out how the LRT nested these two ‘lepidosaurs of uncertain affinities’ two years ago.
In the authors’ Systematic Paleontology section, they reported, “Reptilia Laurenti,1768 Lepidosauromorpha Benton 1983 Squamata Oppel 1811 Genus Oculudentavis Xing et al”
The first two clades are correct. The last two are incorrect. If the reason for the original paper’s retraction was the fact that the original Oculudentavis was incorrectly identified as a dinosaur and a bird, then this paper should likewise suffer the same embarrassment of retraction because neither Oculudentavis specimsn is a squamate nor a lizard. The second specimen is not congeneric with the first Oculudentavis and so needs a new generic name.
It’s the ‘streetlight effect.’ “The streetlight effect is a type of observational bias that occurs when people only search for something where it is easiest to look.”
Bolet et al borrowed four cladograms. All four suffered from taxon exclusion. We’ve seen this sort of error before. It happens all the time in pterosaur studies. Here’s the solution: Build your own LRT with a wide gamut of taxa. Thereafter no tested taxa will ever be of “uncertain affinities.”
The top six problems in paleontology: Number one: taxon exclusion (= the streetlight effect). Number two: borrowing untested cladograms. Number three: trusting genomic results. Number four: trusting textbooks and academic traditions. Number five: freehand reconstructions. Number six: “Pulling a Larry Martin” = focusing on one to a dozen traits, rather than a complete suite. Solution: Keep adding taxa to your own trait-based cladogram. Trace specimens with transparent colors and from those tracings create more accurate and verifiable reconstructions.
References Bolet A, Stanley EL Daza JD, Arias JS ,Cernansky A ,Vidal-Garcı´a M, Bauer AM, Bevitt J, Peretti A and Evans SE 2022. Unusual morphology in the mid-Cretaceous lizard Oculudentavis. Current Biology 31:3303–3314,
Some placoderms are not like the others. Some derived members of the Petalichthyidae (Fig 1), like Late Silurian Qilinyu, are traditional ‘incertae sedis’ placoderms. Wuttagoonaspis (Fig 1) is a traditional arthrodire placoderm. Still others, like Lunaspis and Diandongpetalichthys (also Fig 1) are traditional petalichthyid placoderms. All arise from Metaspriggina, Drepanaspis, Drepanaspis, Arandaspis and Poraspis in the large reptile tree (LRT, 2128 taxa).
Petalichthyidae developed fins by convergence, but remained jawless. All were slow-moving, armored bottom feeders. In this way petalichthyids were convergent with coeval trilobites and extant horseshoe crabs.
Figure 1. Basal jawless placoderms to scale, including Metaspriggina, Arandaspis, Poraspis, and Lunaspis. Increasingly large, orbits rotate dorsally, pectoral fins develop.
Lunaspis heroldi (Broili 1929; latest Early Devonian, 405mya) is traditionally considered a petalichthyid placoderm. The tail, missing from this fossil, was likely long and whip-like, as in rays. None of the specimens preserve mouth parts. Note the serrated leading edge of the pectoral fins.
Figure 2. Subset of the LRT focusing on the clade Placodermi.
Diandongpetalichthys liaojiaoshanensis (Zhu et al. 1991; Early Devonian) is a petalichthid placoderm close to Lunaspis. The postorbital is restricted to the lower orbit. Jaws are lacking. It is considered primitive.
Wuttagoonaspis fletcheri (Ritchie 1973; Middle Devonian) was originally considered ‘an unusual arthrodire’ from Australia. Here it nests with Lunaspis. Note the posteriorly set orbits. The mouth and nares must be ventrally because they are not visible dorsally.
Qilinyu rostrata (Zhu et al. 2016; Late Silurian, 419mya) This mid-sized bottom feeder has a pointed rostrum and a small face. The ventral nostrils are located in front of the weak ventral oral opening. The armor extended to the tiny pelvic fins.
Figure 3. Hemicyclaspis and a variety of other osteostracans in dorsal view created by Ferrón et al. 2020 to test their hydrodynamics.
The other clade of non-placoderm pre-gnathostome fish started off as semi-naked lamprey-like open sea swimmers. Then Birkenia, heterostracans and osteostracans developed armor by convergence. That armor disappeared again with Silurian thelodonts and sturgeons leading to sharks, then bony fish, then tetrapods.
Figure 4. Austroptyctodus and Materpiscis to scale and 90% of life size @72dpi. That’s an embryo at right below the panel that terminates the backbone. Related catfish taxa do not have live birth, but lay hundreds to tens of thousands of eggs.
Another clade of placoderms with fewer, smaller skull bones on narrower skulls, the Ptyctodontida, Among these are Austoptyctodus, Materpiscis (Fig 4) and kin. I
References Broili F 1929. S. B. Bayer. Akad. Wiss., 1 Carr RK, Joahnson Z and Ritchie A 2009. The phyllolepid placoderm Cowralepis mclachani: Insights into the evolution of feeding mechanisms in jawed vertebrates. Journal of Morphology 270(7):775–804. Ritchie A 1973. Wuttagoonaspis gen. nov., an unusual arthrodire from the Devonian of Western New South Wales, Australia. Palaeontographica 143:58–72. Zhu M 1991. New information on Diandongpetalichthys (Placodermi: Petalichthyida). Early vertebrates and related problems of evolutionary biology. Beijing: Science Press. 179–192. Zhu et al. 2012. An antiarch placoderm shows that pelvic girdles arose at the root of jawed vertebrates. Biology Letters Palaeontology 8:453–456.
Jaws and fins evolved twice in vertebrates. As we learned earlier, jaws and fins developed by convergence in placoderms (Fig 1) and gnathostomes. Presently, only the large reptile tree (LRT, 1227 taxa, subset Fig 2) recovers this hypothesis of interrelationships. Prior cladograms excluded too many pertinent taxa and did not completely homologize basal vertebrate bones with tetrapod homologies.
Figure 1. The origin of placoderm fins and jaws as currently derived from Drepanaspis, Phylogenetic miniaturization occurs with tiny Bianchengichthys. Colors added here to mark the homology of skull bones with tetrapods. Blue arrows are water exits from the gills. Qilinyu is not directly related to the upper two taxa, but represents a third appearance of fins, but this time without jaws.
Phylogenetic miniaturization during the origin of fins in placoderms Finless, jawless Drepanaspis (Fig 1) measured 30cm long. Tiny Bianchengichthys (Figs 1,3) measured less than 4cm at the genesis of jaws and fins in placoderms. We’ve seen similar phylogenetic miniaturization at the genesis of novel structures many times before in vertebrate evolution. In this case Bianchengichthys evolved weak jaws and tiny fins alongside the convergent appearance of fins in Thelodus and later toothless jaws in Chondrosteus.
Figure 2. Subset of the LRT focusing on placoderms.
Li et al thought Bianchenichthys was far from the origin of jaws and did not realize the placoderm origin of jaws was different from and convergent to the gnathostome origin of jaws. Drepanaspis was not mentioned in their text. Their figure 4 indicated a sister relationship between the Late Silurian placoderm Entelognathus and the Late Devonian lobefin Eusthenopteron. The LRT does not support the Li et al 2022 hypothesis of interrelationship. Without a valid cladogram testing a panorama of taxa, taxon excluded results are suspect.
Figure 3. Bianchengichthys traced and reconstructed using DGS methods. Compare to the freehand reconstruction from Li et al in figure 1. The median line was added and does not represent a suture. The blue-green arrow represents water exiting from the single gill opening behind the jugal.
Drepanaspis gemuendenensis (Schlüter 1887; Gross 1963; Early Devonian 405mya; up to 30cm, Fig 1) was a large, flattened arandaspid with a wde, superficially ray-like armored body. The common gill opening exited posteriorly. This bottom feeder with widely-spaced eyes is traditionally considered a member of the Heterostraci. Here it nests at the base of the Placodermi.
Bianchengichthys micros (Li et al. 2021, Latest Silurian, IVPP V27128A, B). Derived from fin-less, jaw-less Drepanaspis, Bianchengichthys has tiny fins and weak jaws convergent with those developed independently in gnathostomes. The oribit + naris opening is anteriorly oriented, as in Arandaspis (Fig 1), likely by reversal due to neotony. Bianchengichthys was reported to be,“exquisitely preserved with mandible and paired fins. Systematically, the new taxon is close to the last common ancestor of bony and cartilaginous fishes. The mandible interestingly combines characters from arthordires and Entelognathus.” Here Bianchengichthys nests basal to those placoderms and far from the origin of bony and other fish.
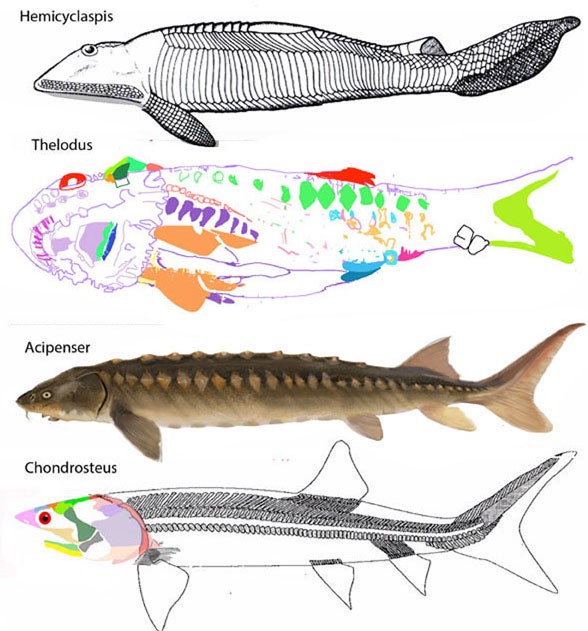
Figure 4. Origin of gnathostome jaws from the ostracoderm, Hemicyclaspis, Thelodus, Acipenser (sturgeon) and Chondrosteus.
Also note Jawless Qilinyu, Wuttagoonaspis and Lunaspis are traditional placoderms that now nest with the Arandaspis – Poraspis clade. So, let’s just call the whole clade, Placodermi (Fig 2).
As reported earlier, gnathostome jaws developed in the pre-shark, post-sturgeon, Chondrosteus (Fig 4), by convergence.
This appears to be a novel hypothesis of interrelationships. If not, please provide a citation so I can promote it here.
References Hu Y, Lu J and Young GC 2017. New findings in a 400 million-year-old Devonian placoderm shed light on jaw structure and function in basal gnathostomes. Nature Scientific Reports 7: 7813 DOI:10.1038/s41598-017-07674-y Li Q et al. 2021. A new Silurian fish close to the common ancestor of modern gnathostomes. Current Biology 31:3613–3620. https://doi.org/10.1016/j.cub.2021.05.053
Today’s post was prompted by running across the above headline and website. It was updated Novermber 8, 2023 after a second origin of jaws split bony fish prior to the origin of bony fish a few days earlier.
Reported by Anietra Hamper in 2019 under the title: “Fish Finds: Uncovering the Origins of the Catfish Family Tree” and the subhead: “How evolutionary biologists are using nearly 50-million-year-old fossils for insight into the origins and evolution of catfish.”
The featured taxon was Middle Eocene Hypsidoris farsonensis (Lundberg and Case 1970, Figs 1, 2), which was subsequently added to the large reptile tree (LRT, 1227 taxa). Unlike many catfish, Hypsidoris retains a toothless bit of loose maxilla.
Figure 1. Hypsidoris in situ. Average length = 20cm.
The featured expert was Dr Lance Grande, Field Museum, Chicago, USA, who redescribed Hypsidoris in 1986.
Figure 2. Closeup of Hypsidoris skull. Colors and restoration added here. Maxilla is green and lacks teeth.
Hamper et al. 2019 reported, “The telltale characteristics of catfish – peculiar denticular pectoral and dorsal spines, the lack of scales on the body, cat-like whiskers, the lack of maxiilary teeth in most species, and exceptional sensory ability, fascinate evolutionary biologists as much as they do the passionate anglers who are drawn to the living versions of catfish today.”
Be careful. Taxa are defined by their phylogenetic ancestors, not their traits.
“The only other catfishes that have a well-developed maxilla with teeth today are species in the South American family, Diplomystidae.”
With that bit of info, Diplomystes (Figs 3, 4) was likewise added to the LRT.
Figure 3. Diplomystes in vivo.
Teeth on the maxilla sounds like it should be primitive. That makes sense, but it’s “Pulling a Larry Martin.“
Figure 1. The catfish Diplomystes has a large branchial basket (pink). Each pair holds a gill.
Adding taxa nests catfish with Guiyu, Dialipina and Loganellia among Silurian taxa.
Figure 5. Late Silurian Guiyu with catfish homologs. See Clarias in figure 2 for comparison.
According to the LRT (Fig 5) the toothy catfish, Diplomystes (Figs 3,4) is not the most primitive catfish. Rather, with the presently tested list, toothless, maxilla-less catfish that more closely resemble Clarias are more primitive. The maxilla of catfish is not homologous with the maxilla of tetrapod ancestors going back to Lasanius and Birkenia.
It is possible that teeth re-evolved in toothy catfish. More likely: catfish had a long ghost lineage back to the Late Devonian not currently represented in the fossil record. Those catfish clades radiated in the meantime.
Figure 8. Hoplosternum in vivo. You can see the armor/bone beneath its shiny skin. Overall it looks more like (= shares more traits with) Robustchthys (Figs 6,7) than toothy catfish (Figs 3,4).
The LRT documents the ancestry of all included taxa back to Ediacaran worms. Adding taxa resolves all phylogenetic problems.
If you go to university you will learn exactly what they teach out of an approved, but outdated textbook.
Instead, if you want to learn as much as you can about evolution, do the work. Build your own LRT. Learn the way things really are and were. Workers won’t like your independence, and you may be ostracized, but you’ll sooner or later understand what they don’t and won’t teach at the university level.
The top six problems in paleontology: Number one: taxon exclusion. Number two: borrowing untested cladograms. Number three: trusting genomic results. Number four: trusting textbooks and academic traditions. Number five: freehand reconstructions. Number six: “Pulling a Larry Martin” = focusing on one to a dozen traits, rather than a complete suite. Solution: Keep adding taxa to your own trait-based cladogram. Trace specimens with transparent colors and from those tracings create more accurate and verifiable reconstructions.
References Grande L 1986. Redescription of Hypsidoris farsonensis (Teleostei: Siluriformes), with a reassessment of its phylogenetic relationships. Journal of Vertebrate Paleontology 7:24–54. Lundberg JG and Case GR 1970.A New Catfish from the Eocene Green River Formation, Wyoming. Journal of Paleontology. 44 (3): 452. Xu G-H 2019. Osteology and phylogeny of Robustichthys luopingensis, the largest holostean fish in the Middle Triassic. PeerJ doi: 10.7717/peerj.7184. eCollection 2019.
Riguetti, Apesteguía and Pereda‑Suberbiola 2022 have put together the bits and pieces of a Mid-Late Cretaceous (95mya) bipedal armored phytodinosaur, Jakapil kaniukura, MPCA-PV-630 (Fig 1). The authors restored most of the bones of Jakapil based on phylogenetic bracketing. Their cladogram nested their find with two quadrupedal early Jurassic armored dinosaurs, Scutellosaurus and Scelidosaurus (Fig 2) and a third pre-stegosaur, Isaberrysaura, all lacking the strong predentary found in Jakapil.
Figure 1. What little is known of Mid-Cretaceous Jakapil compared to the more complete and much younger Late Jurassic Yinlong. Note the large predentary and extremely deep mid-mandible. Yinlong may be a better fit for Jakapil, but Jakapil will not be tested in the LRT due to a paucity of material.
Traditionally the armored dinosaurs = stegosaurs + anklyosaurs, are considered members of the clade Thyreophora.
Unfortunately, the large reptile tree (LRT, 2125 taxa, Fig 3) does not recognize Thyreophora as a monophyletic clade, unless it also includes hadrosaurs and ceratopsians. According to the LRT, stegosaurs and ankylosaurus evolved armor by convergence, like another phytodinosaur with armor, the sauropod, Saltasaurus.
Figure 2. Scelidosaurus, Isaberrysaura, Yinlong, Jakapil and Scutellosaurus to scale. Yinlong is the best match to Jakapil here, but is the outlier in the Jakapil paper.
According to Wikipedia, “Jakapil represents a novel morphotype among thyreophorans, including, among other things, the presence of a predentary bone (absent or cartilaginous in other basal thyreophorans[), large, low osteoderms, and a bipedal stance. Its describers estimate it to be 1.5 metres (4.9 ft) long and 4–7 kilograms (8.8–15.4 lb) in weight, based on femoral circumference.”
Perhaps not that novel. After all, the authors nested Jakapil with other armored dinos. Even so, maybe there’s a closer match among the bipedal ceratopsia (Fig 1) sans scutes.
Figure 3. Subset of the LRT focusing on Ornithischia. Traditional Thyreophora include only the blue and violet clades, but note: Yinlong is a descendant.
The extremely deep mid-mandible of Jakapil is a distinct character. A precursor deep mid-mandible can be found in the earlier basal ceratopsian, Yinlong (Fig 1) from the Late Jurassic of China. And it has a strong predentary. Like Jakapil,Yinlong is a biped with a similar, small, hockey-stick scapula and small forelimbs not seen in recovered sister taxa (Fig 2). Yinlong was tested by the authors, but it lacks bony scutes. In the LRT (Fig 3) Scelidosaurus was in the ancestry of Yinlong. Given what little is preserved of Jakapil (Fig 1) perhaps more weight was put on the scute traits during original scoring. Too few bones are known from the Jakapil holotype to attempt to nest it in the LRT.
References Riguetti FJ, Apesteguía S and Pereda‑Suberbiola X 2022. A new Cretaceous thyreophoran from Patagonia supports a South American lineage of armoured dinosaurs. Nature Scientific Reports 12:11621
Carr et al 2022 listed six problems with Paul et al 2022, criticizing that earlier and widely publicized lumping and splitting of the genus Tyrannosaurus (Fig 1). Read the Carr et al list and see if you can figure out how both sides got published without a key ingredient.
Carr et al reported: 1. “the taxon diagnoses are based on two features that overlap between the species;” 2. “several skulls cannot be identified based on the diagnoses; “ 3. “proportional comparisons between Tyrannosaurus and other theropods are based on incomparable samples;” 4. “the tooth data are problematic;” 5. “the stratigraphic framework divides the Hell Creek Formation into thirds, without the stratigraphic position of each specimen, or independent age control showing the subdivisions are coeval over the entire geographic area;” 6. “previous work found variation in T. rex, but it cannot be parsed into discrete categories.”
Figure 1. Tyrannosaurus specimens under study. Images from Paul et al 2022. Colors added.
Note that neither Carr et al 2022 nor Paul et al 2022 performed a phylogenetic analysis using several hundred traits on several specimens of Tyrannosaurus and their outgroups [but see PS addendum below]. Analysis remains the best and only way to correctly lump and split taxa. Cherry-picking a few traits, like Paul et al did and Carr et al argued against, represents “Pulling a Larry Martin.” If the Tyrannosaurus specimens can be split into discrete clades or not is the only question. Only analysis will answer that question. Both sets of workers have been caught dancing around this issue without diving into it. Apparently neither team wants to do the necessary work. Here’s what they needed to do: Perform the analysis. Both sides should do this, sharing all data. Then these Tyrannosaurus analyses can be tested for confirmation, refutation or correction. Afterwards there should be no argument because they should both come to the same unbiased conclusion.
Is paleontology a zero-sum activity? (= a situation in which whatever is gained by one side is lost by the other). I’ve wondered this for a long time. Carr et al. didn’t want Paul et al. to succeed in splitting Tyrannosaurus. They felt moral outrage. If anyone were to ever split Tyrannosaurus, it should be Carr et al. Paul has no degree in paleontology. He and his team reported on their studies.
On a similar note, when I discovered four proximal ancestors of pterosaurs in Peters 2000 using three different phylogenetic analyses, pterosaur workers respond by saying ‘we prefer other taxa without testing yours‘. Some resorted to name-calling. Evidently they also saw paleontology as a zero-sum activity. By reporting on the ancestors of pterosaurs that was one more discovery they could never discover because it had been plucked off the branch by someone else. Evidently they thought I took something that belonged to them. They had degrees. They taught at the university level. I was a rank amateur with no expenses. No wonder I was blackballed. Those workers had and continue to have moral outrage. Historically that’s typical. Still not forgivable. Scientists are human and they, like everyone reading this, are ruled by their limbic system. That needs to change, if possible.
On a quasi-similar note, my rejected manuscript for “The triple origin of whales, currently available on ResearchGate.net has had 1000+ reads as of last week. That means that many workers were curious enough to download the PDF, but were not brave enough to talk about that banned study out in the open. At least no one in the profession has written to me to offer advice, guidance or editing on that manuscript. Why? Because paleontology is a zero-sum activity in their minds. Evidently no one wants to be told their genomic studies were useless to harmful. No one wants to be told they missed including a few pertinent taxa.
If you’re an aspiring paleontologist, and you encounter this sort of reaction and rejection, relax, it’s common, historical and widespread. Unlike astronomers, paleontologists think paleontology is a zero-sum activity. Not sure how to change that mindset.
PS This same afternoon a reader alerted me to an earlier Tyrannosaurus cladogram from Carr 2020 (Fig. 2).
Figure 2. Tyrannosaurus cladogram from Carr 2020. Colors added here to link specimens reported by Paul et al 2022. Note there is no pattern here that matches the few species split of Paul et al.. However, among the most derived taxa the regina species clade does nest apart from the rex and imperator species clades.
The Carr 2020 cladogram matches the large reptile tree (LRT, 2125 taxa) in nesting USNM 555000 (=T regina orange) outside of the CM 9380 (=T rex green) + FMNH PR 2081 (T imperator cyan) clade. The outlier, the RSM 2523.8 specimen (green), was reported by Carr 2020 as “one of the least mature adults is also the largest and most massive example of the species.” According to the Carr cladogram, if the Tyrannosaurus genus is split, the RSM specimen will certainly need its own species.
Figure 3. Subset of the LRT focusing on Tyrannosaurus.
Carr 2020 also reports, “The extreme number of changes at the transition between juveniles and subadults shows that the ontogeny of T. rex exhibits secondary metamorphosis, analogous to the abrupt ontogenetic changes that are seen at sexual maturity among teleosts.”
Scale illustrations of pertinent specimens with overlapping traits preserved would be helpful here. Otherwise we can anticipate the lumpers and splitters will both stick to their original hypotheses. They both have a case according to the cladograms.
And finally… three posts ago we passed the 4000 posts landmark. It’s been a good run.
References Carr TD 2020. A high-resolution growth series of Tyrannosaurus rex obtained from multiple lines of evidence. PeerJ 8:e9192 https://doi.org/10.7717/peerj.9192 Carr TD et al. (6 co-authors) 2022. Insufficient Evidence for Multiple Species of Tyrannosaurus in the Latest Cretaceous of North America: A Comment on “The Tyrant Lizard King, Queen and Emperor: Multiple Lines of Morphological and Stratigraphic Evidence Support Subtle Evolution and Probable Speciation Within the North American Genus Tyrannosaurus.”. Evolutionary Biology https://doi.org/10.1007/s11692-022-09573-1 Paul GS, Persons WS and Van Raalte J 2022. The tyrant lizard king, queen, and emperor: Multiple lines of morphological and stratigraphic evidence support subtle evolution and probable speciation within the North American genus Tyrannosaurus. Evolutionary Biology. https:// doi. org/ 10. 1007/ s11692- 022- 09561-S