Habib and Pittman 2021 report:
“The flight capabilities of Archaeopteryx continue to be highly contentious. We addressed locomotor performance in this iconic animal using ratios of forelimb to hind limb bone cross-sectional properties.”
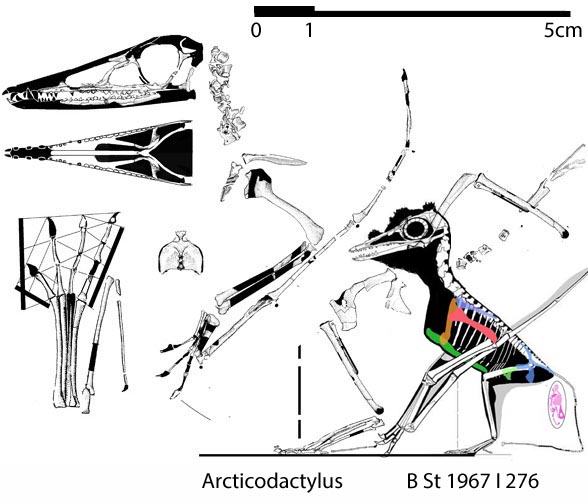
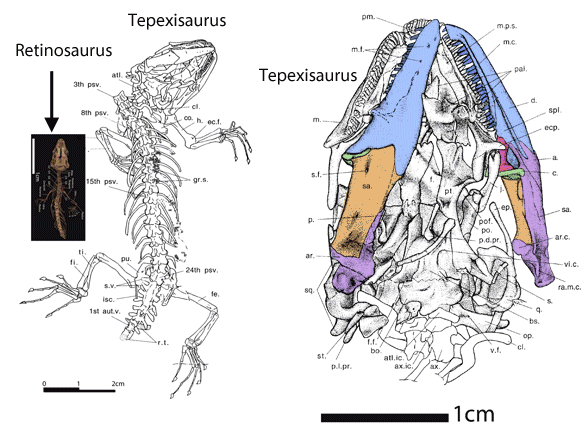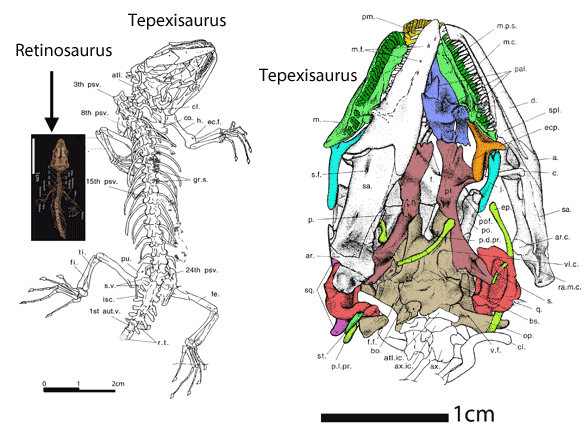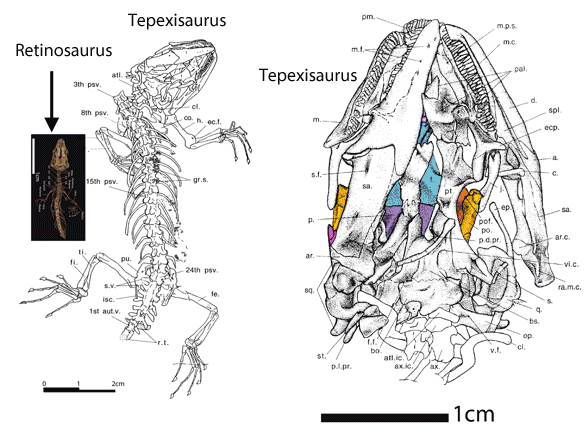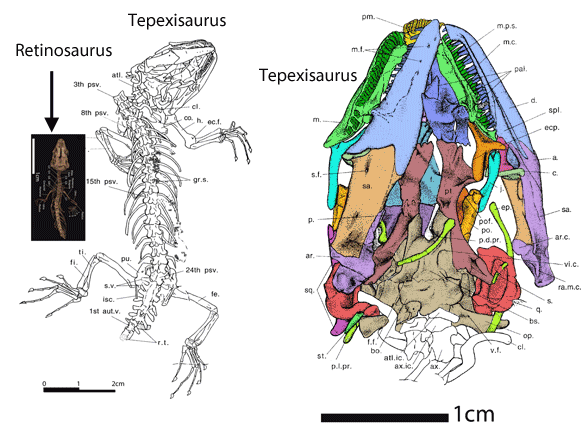
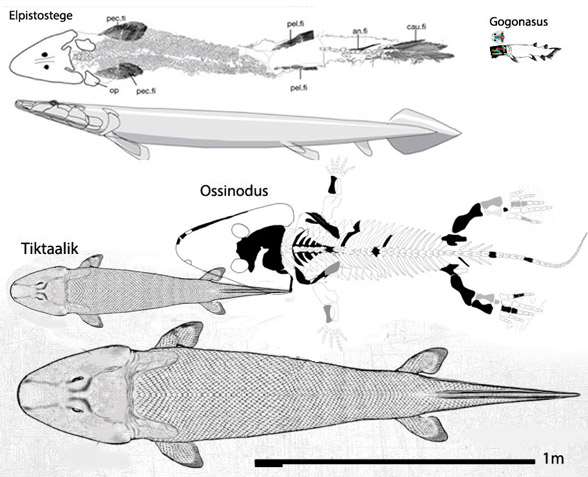
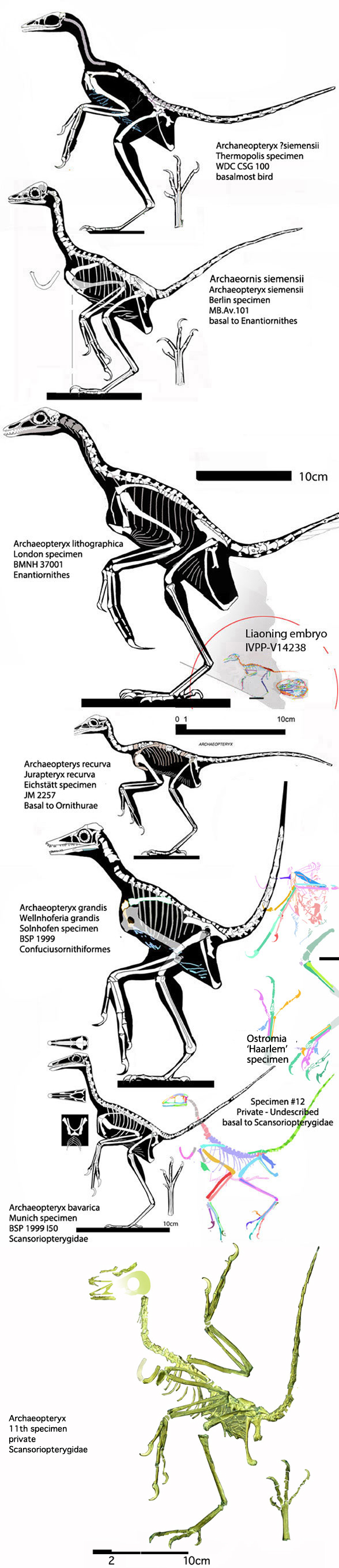
Which one of the 13+ Solnhofen birds did Habib and Pittman look at? Every fossil is different (Fig. 1). PhDs need to address these differences and not employ nebulous wastebasket nomenclature.
Habib and Pittman continue:
“We compared these results to the same ratios in living birds and the dromaeosaurid, Rahonavis, which provided a critical paravian out-group.”
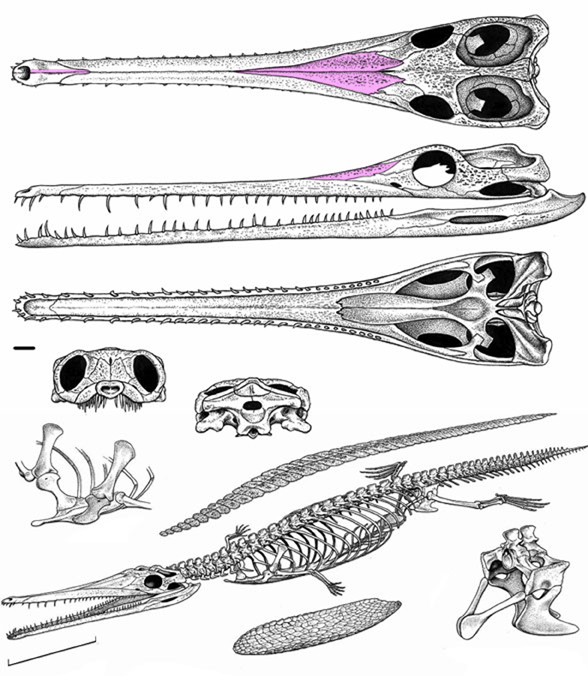
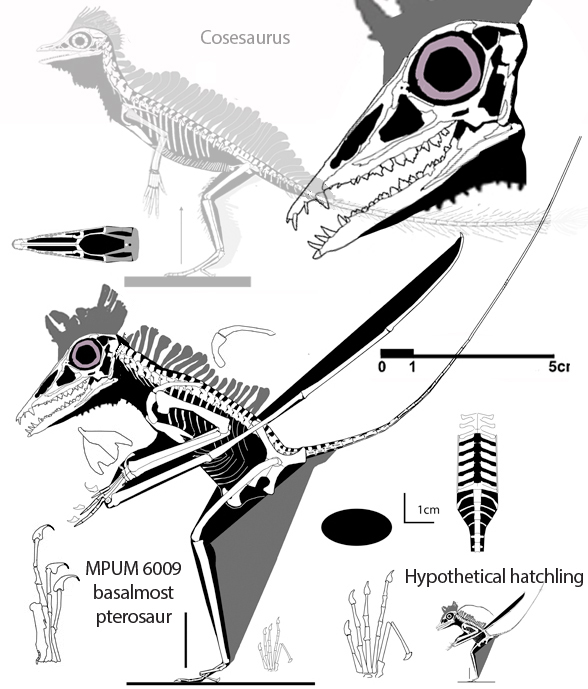
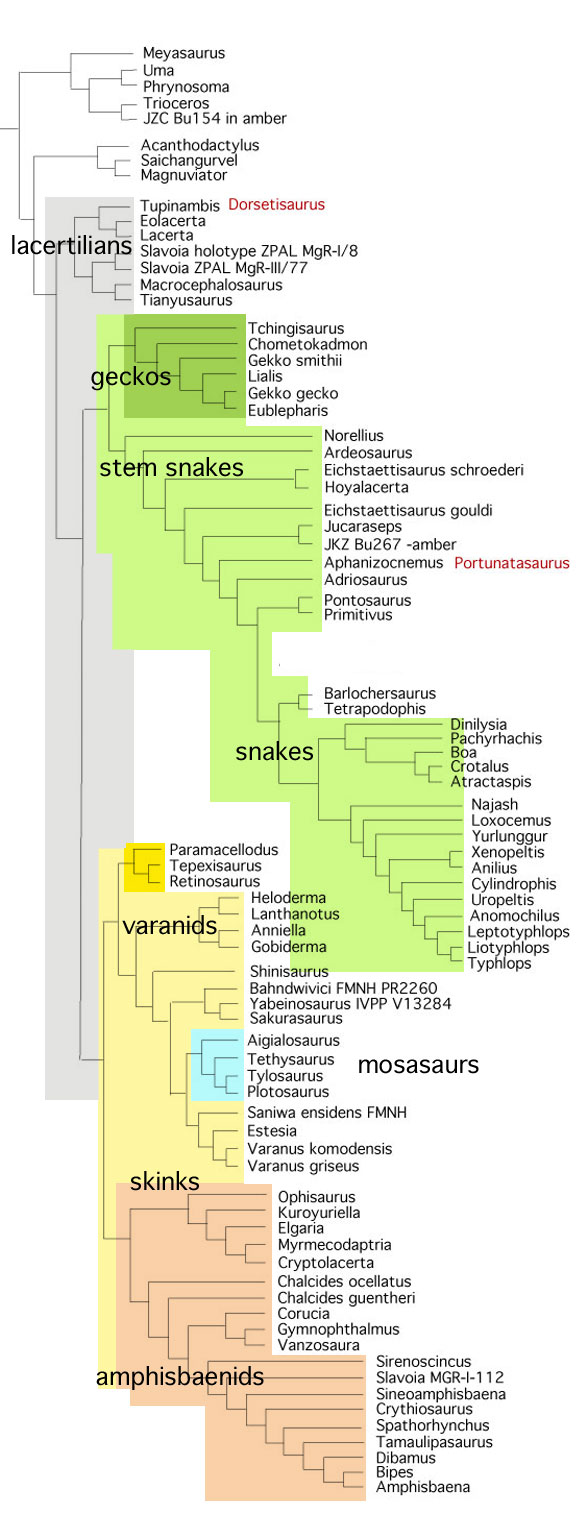
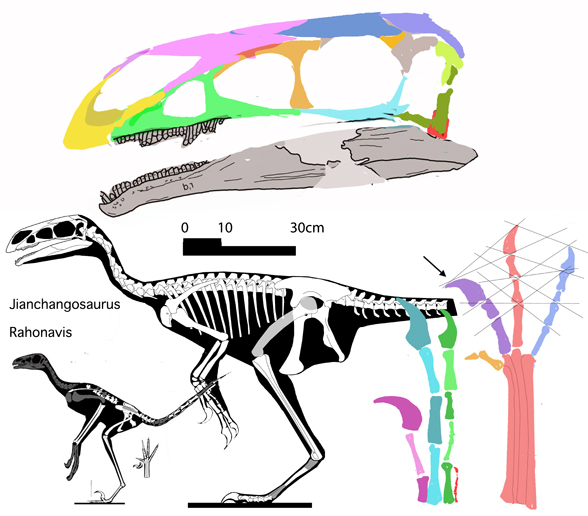
In the large reptile tree poorly-represented, Rahonavis (Fig. 2), is a small Late Cretaceous (70mya) therizinosaur close to Jianchangosaurus (Fig. 2) from the land of oddball taxa, Madagascar. Both of these therizinosaurs look superficially like dromaeosaurids.
Both have a big toe two ungual.
The problem is: neither are paravian out-group taxa. So, what were these two PhDs thinking when they cherry-picked an incomplete Late Cretaceous taxon and said it was ‘a critical paravian out-group’ of a Late Jurassic radiation?
Why not choose a valid and coeval out-group taxon like Almas? Sinovenator? Anchiornis? The Daiting specimen attributed to Archaeopteryx? Xiaotingia? Ostromia? Eosinopteryx?
Or all of them?
Habib and Pittman continue:
“Our analysis recovered Rahonavis as a capable powered flyer.”
No relatives of Rahonavis fly. Only the radius and ulna are preserved in Rahonavis (Fig. 2), not the entire forelimb. Habib and Pittman are guessing what the humerus and manus looked like.
Advice: Choose a more complete and coeval taxon to work with.
A little backstory: Co-author Habib invented the quad-launch in pterosaurs hypothesis by cheating the morphology to suite his vision. Habib also co-authored a soaring azhdarchid hypothesis (Witton and Habib 2010) to suite his vision. This latest abstract does not repair his reputation as Habib continues to overlook inconvenient facts.
“Conversely, we found Archaeopteryx to be a less proficient long-distance flyer than Rahonavis, despite being closer to crown group birds.”
Conclusions like this, based on incomplete data, should be ignored.
“Archaeopteryx was highly cursorial and a short-ranged flyer, at most. The scaled humeral/ femoral section modulus ratio in Rahonavis was 1.49, more than four times higher than the ratio of 0.32 recovered for Archaeopteryx.”
The humerus is not known for Rahonavis (Fig. 2). Neither is the manus. The authors guessed based on imagination and produced a conclusion on that guess. Cherry-picking like this invites criticism. Habib and Pittman could have avoided criticism by using coeval pre-bird taxa, but they ignored published cladograms.
BTW, Rahonavis was also ‘higly cursorial’. Just look at those long slender legs.
“The bone strength ratios in Rahonavis are close to (within 99% Confidence Interval) those measured for falcons, pheasants, and egrets: birds that have strong hind limbs but retain excellent flight abilities (sustained in falcons and egrets, short-range bursts in pheasants).”
Bone strength ratios again. Habib used similar statistics and bone strength ratios to introduce the pterosaur quad-launch hypothesis. That’s fine so long as you don’t short-change or cheat morphology and avoid systematics.
Advice: Dr. Habib and Dr. Pittman, please use a cladogram next time. Don’t cherry-pick invalid outgroup taxa separated in time by tens of millions of years and omit closely related taxa.
Maybe Rahonavis was a capable flyer by convergence. Maybe not. Always start with a valid phylogeny to avoid problems like these two authors have dug themselves into.
“Archaeopteryx compares closely in its humeral/femoral strength ratios with secondarily flightless cursorial birds (99% CI).”
That is to be expected as Solnhofen birds were just learning to fly.
Again, Habib and Pittman need to show that phylogenetic precursors were more flight capable. And which Archaeopteryx (Fig. 1) did they test? Did they pick one to suit their narrative?
“It seems to have possessed a combination of femora that were moderately stronger than expected for its size and humeri that were much more gracile than predicted from regressions of section modulus against body mass for extant birds. Our results are indicative of secondary flight reduction or loss in Archaeopteryx, especially when combined with the existing literature data on this legendary taxon.”
Solnhofen birds became legends because 1) they were considered the first theropods capable of flight; 2) they were considered the first dinosaurs with feathers. Comparing one Late Jurassic Solnhofen bird to one incomplete Late Cretaceous (70mya) therizinosaur does not indicate ‘secondary flight reduction or loss in Archaeopteryx.’
“In a secondary flight reduction model, seemingly conflicting results of prior research regarding the flight capabilities of Archaeopteryx are explained as variably detecting volant ancestry versus flightless life history.”
Habib and Pittman are painting a picture with taxa that are not related to one another, either phylogenetically or chronologically.
“We suggest that Archaeopteryx might have had an ecology and locomotor profile like that of living island rails, many of which are secondarily flightless or flight-reduced hunters that use a wide range of wing-assisted behaviors.”
That’s not a novel suggestion.
“Archaeopteryx may have been a similarly capable island hunter, relying on its exceptionally long and robust hind limbs to chase down small animals and its reduced wings to assist with especially acrobatic terrestrial maneuvers. Our new picture for Archaeopteryx is not one of an animal that was “suboptimal” in any respect. Instead, we reconstruct Archaeopteryx as a specialized, fast, and agile island endemic.”
First straighten out the phylogeny, then the chronology, then avoid the bad habit of cherry-picking incomplete taxa far too late in time in order to suit a narrative. Instead: build a cladogram. Use valid outgroup taxa. Draw conclusions from those.
References
Habib MB and Pittman M 2021. Going up or coming down? Archaeopteryx as the earliest known secondarily flight-reduced vertebrate. Journal of Vertebrate Paleontology abstracts.
Witton MP and Habib MB 2010. On the Size and Flight Diversity of Giant Pterosaurs, the Use of Birds as Pterosaur Analogues and Comments on Pterosaur Flightlessness. PLoS ONE 5(11): e13982. https://doi.org/10.1371/journal.pone.0013982
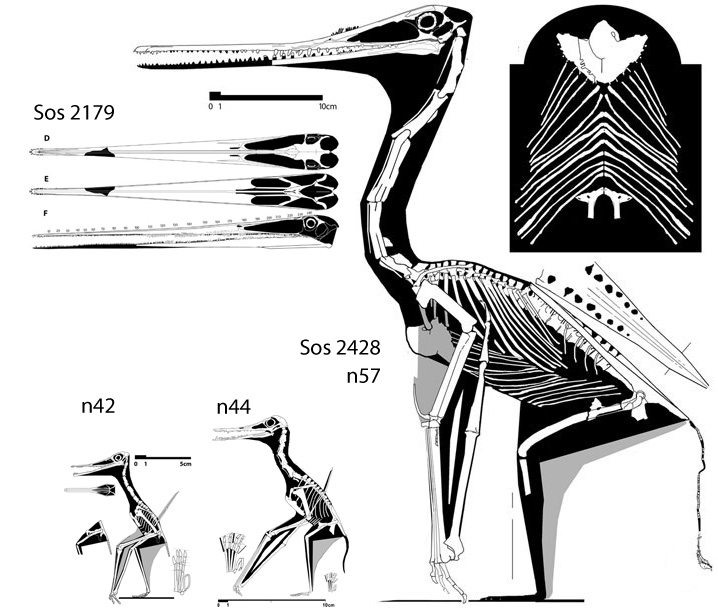
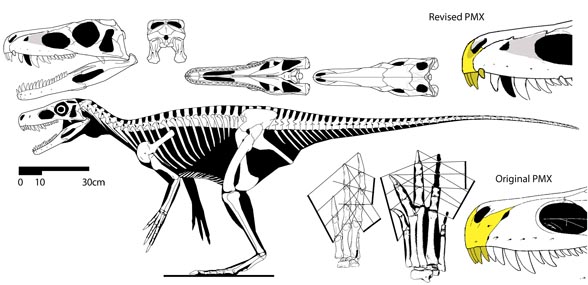
‘Earliest secondarily flight-reduced vertebrate?‘ No.
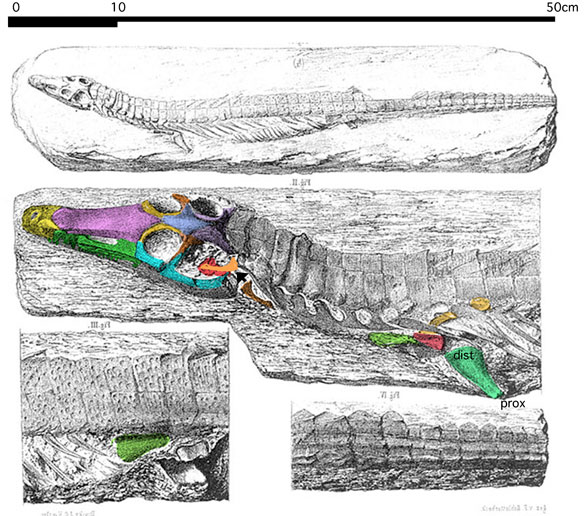
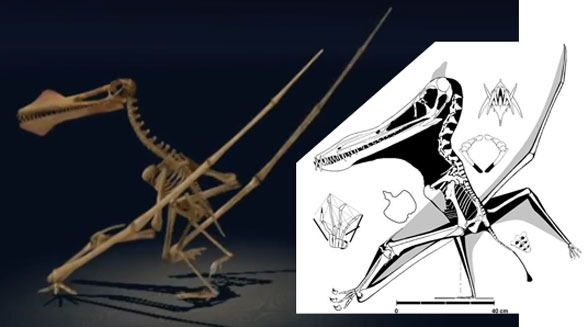
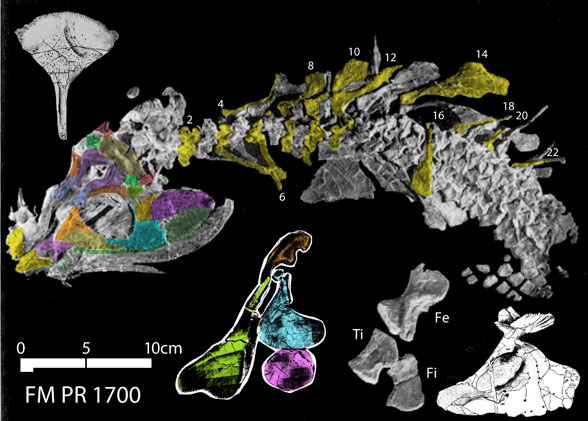
That accolade goes to the flightless pterosaur, SOS 2428 (Fig. 3), also from Solnhofen limestones, chronicled here and at Researchgate.net.