Just wanted to take a moment to remember Dr. Hal Levin, professor emeritus, Earth and Planetary Sciences, Washington University, St. Louis, USA. He entered our profession in 1961, became department chair in 1973 and died in 2019 at age 90. He was the author and often illustrator for eleven editions of his geology textbook, “The Earth Through Time.”
Fig 1. Hal Levin, Wash U professor and author
From Levin’s obituary: “Throughout his life, Levin was committed to unraveling the Earth’s secrets by examining microscopic ancient invertebrates from the Earth’s sedimentary layers to predict future changes and to expose the deleterious effects of climate change.
“Levin joined the Washington University faculty in 1961. He taught and conducted research while concurrently serving as the coordinator of preprofessional studies in Arts & Sciences.
“He was chair of the Department of Earth and Planetary Sciences from 1973-76. A prolific author, Levin wrote and illustrated numerous textbooks and laboratory studies in geology and paleontology, including 11 editions of the popular geology textbook “The Earth Through Time.”
Back in the day when I was illustrating books like ‘Giants‘ …and back when you could just walk into university libraries and offices without a digital ID and a face mask, Dr. Levin invited me to illustrate the cover for the fifth edition of his university-level textbook on geology, ‘The Earth Through Time’ (Fig 2). I was just getting started in paleontology back then. He was just getting ready for retirement.
Figure 2. ‘The Earth Through Time’ the fifth of eleven editions, by Harold (Hal)( Levin, WashU. Cover illustration by David Peters.
Wash U is not famous for fossils, but has several impressive legacy specimens from the old Gustav Hambach collection. Habach was a German immigrant who became head of the geology dept in 1887. Since then Wash U has been more into geology, both here on Earth and on Mars, with forays into human origins during the heyday of David Tab Rasmussen and Glenn C Conroy.
Figure 3. Ichthyosaur from the Hambach collection, Washington U, St Louis, USA.
Hal Levin was always kind and accommodating, perhaps because there have never been more than a few paleo professionals and enthusiasts in St. Louis. Somehow we all find our mentors.
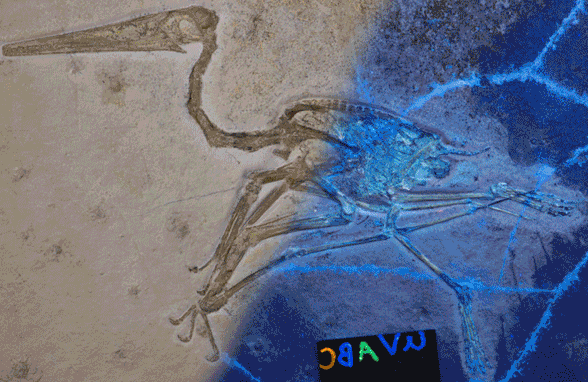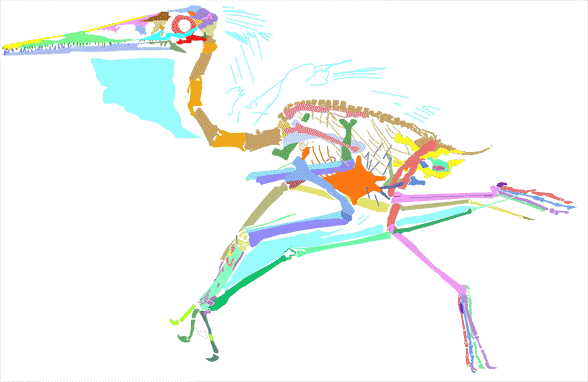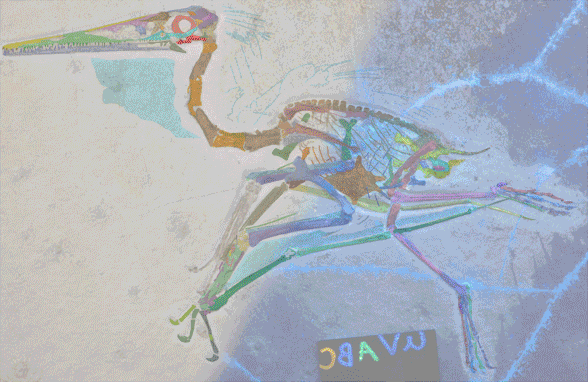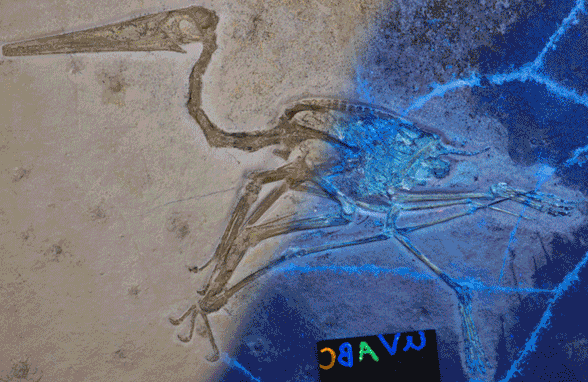
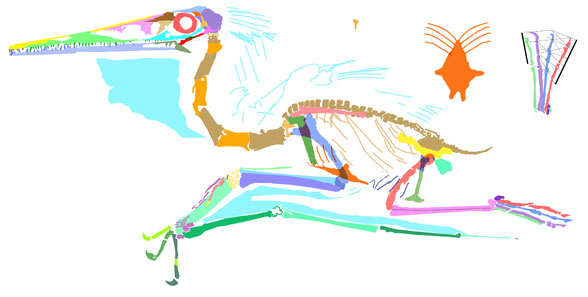
This new Pterodactylusspecimen (Figs 1, 2, LF 101, 8cm skull length) came to my attention from a 2020 online Houston Museum of Natural History story featuring photographer Mike Ecklund. He sometimes uses ultraviolet (UV) light frequencies in an attempt to tease out more data.
Figure 1. Pterodactylus 2 from the Lauer collection, flipped left to right. 1. As originally published on the HMNS website with half of the image in UV light. 2. Traced in colors (DGS). Note the soft tissues.
“The naked eye doesn’t tell enough of the story.” Eklund reminds us. That’s true, but let’s dive deeper into the story. Sometimes it takes digital graphic segregation (DGS) to help the naked eye focus on every millimeter of the skeleton and ultimately step back to create a reconstruction which can be scored in analysis.
Figure 2. Pterodactylus 2 from the Lauer collection reconstructed from DGS tracings in figure 1. Pterosaur workers have been averse to adopting this method, first emailed to them in 2003.
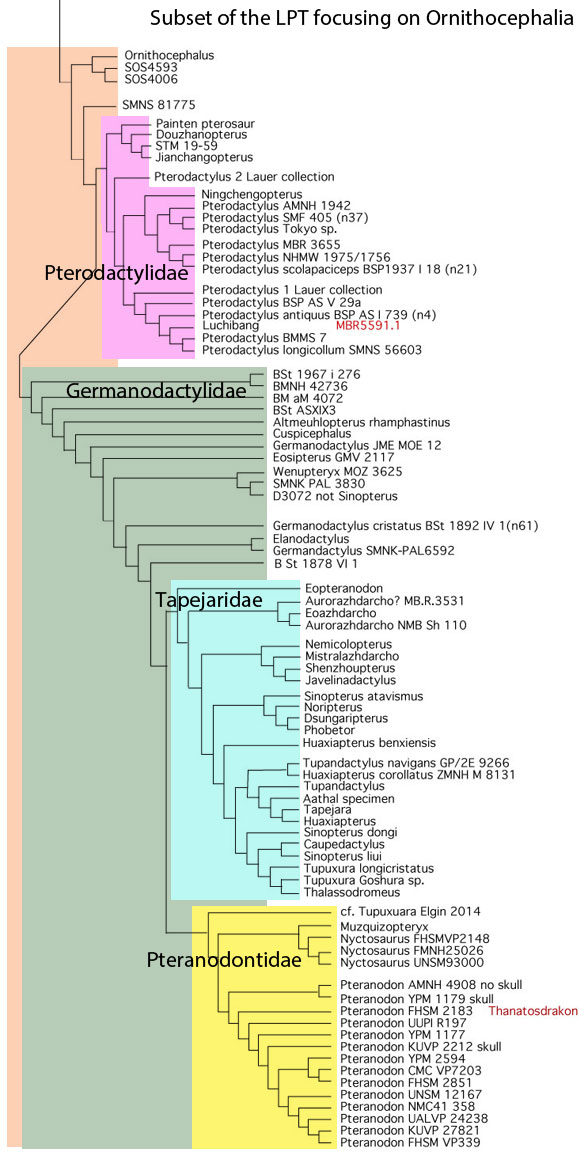
When Pterodactylus 2 Lauer collection was added to the large pterosaur tree (LPT, 261 taxa, subset Fig 3) it nested basal to most Pterodactylus specimens. The specimen (Fig 1) is largely articulated and preserves soft tissue. A reconstruction (Fig 2) shows the specimen in repose. The skull preserves a naris separate from the antorbital fenestra, manual digit 5, a wing ungual, a shallow chord wing membrane, a gular sac and faint traces of dorsal extradermal pycnofibers.
Figure 3. Subset of the LPT focusing on Ornithocephalia, the hummingbird-sized ancestors of Pterodactylus, Germanodactylus, Taepara and Pteranodon.
Recent housekeeping in the LPT (subset Fig 3) removed three taxa known from scrappy skeletal material (now listed in red), and rescored several other taxa first added a decade ago. This resulted in a cladogram with complete resolution, except at one node at the base of the genus Pteranodon where skull-only specimens nest with post-skull-only specimens. The only topology change moved Eopteranodon and kin from the base of Pteranodontidae one node over to the base of the Tapejaridae.
If you note any major topology differences between the LPT and the much smaller cladograms produced by other pterosaur workers, those are due to taxon exclusion. Other workers refuse to add long lists of Dorygnathus and Scaphognathus taxa. Fifteen years ago Peters 2007 indicated such additions revise cladograms, splitting the traditional clade ‘Pterodactyloidea’ into four convergent pterodactyloid grades. As you can tell from their response, that bit of insight has not been welcome news to those who have been teaching about ‘pterodactyloids’ for the past decade and a half.
This lack of curiosity and testing among pterosaur PhDs should not be generalized to PhDs in other sciences. Pterosaur workers are a unique clade averse to adding taxa and straying from Benton’s textbook, from which they earn their living.
YouTube video featuring photographer Mick Ecklund
References Abel O 1925. On a skeleton of Pterodactylus antiquus from the lithographic shales of Bavaria, with remains of skin and musculature. American Museum Novitates 192: 12 pp. Peters D 2007. The origin and radiation of the Pterosauria. In D. Hone ed. Flugsaurier. The Wellnhofer pterosaur meeting, 2007, Munich, Germany. p. 27. Wellnhofer P 1970. Die Pterodactyloidea (Pterosauria) der Oberjura-Plattenkalke Süddeutschlands. Abhandlungen der Bayerischen Akademie der Wissenschaften, N.F., Munich 141: 1-133.
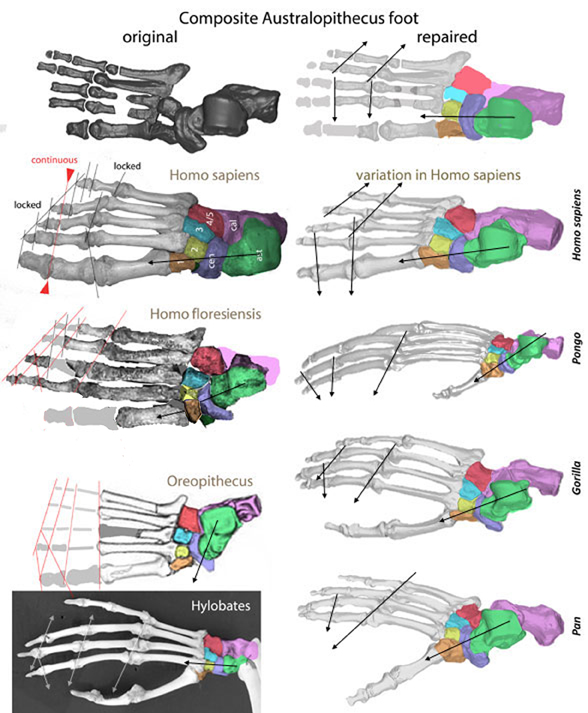
Earlier we looked at several ape pedal (= foot) bones in an effort to understand changes between terrestrial- and arboreal-adapted bauplans with the question: which one of these candidates is closest to humans?
Today this thread continues with more included taxa, including a second, more gracile Homo sapiens (Fig 1 right column) example showing variation distinct from the original robust one (in the left column).
Some PILs (parallel interphalangeal lines, Peters 2000a) are shown to assess their flexibility (with continuous PILs) or inflexibility (with discontinuous PILs).
Figure 1. Selection of several ape pedes. Colors added here. Note the variation with the two examples of Homo sapiens. The Australopithecus pes was repaired by moving distal carpal 3 to distal carpal 2 and flipping the centralia so it contacts carpal 3 in accord with the other pedes.
Metatarsal 2 alignment In the left column mt2 is deeper than mt3. In the right column it is not deeper than mt3 with the possible exception of Gorilla. Alignment permits flexion. Misalignment prevents flexion at the joint series. In all these apes mt2 is deeper than mt1.
Pedal digit 1 axial rotation Most of the apes (Fig 1) have their ventral side in contact with the substrate. Hylobatesand Gorilla are exceptions with digit 1 axially rotated with the ventral side oriented medially. The Homo pes in the left column also shows a bit of axial rotation, not duplicated in the Homo pes in the right column. More importantly, note in Pan and Gorilla, digits 2–4 are exposing their medial surfaces. That’s just another way of achieving the same morphology. In any case, this rotation contributes to a transverse arch in the sole of the foot, distinct from most tetrapods.
Tarsus variation In most examples (Fig 1) the angle between the astragaulus (green) and centralia (lavender) has a medial vector extending to a medially directed digit 1. Hylobates and Homo have a more anteriorly directed vector running through digit 2.
Tarsus size In most examples (Fig 1) the tarsus is relatively robust (>40% of the pedal length). In the arboreal Hylobates and Pongo the tarsus is much reduced relative to the digits and metatarsals.
First set of interphalangeal joints In Pongo and Pan this first PIL extends beyond pedal digit 1. In Gorilla this PIL extends to pedal 2.2. In the other genera this PIL extends to pedal 2.1 or to the base of pedal 2.1 (the metatarsophalangeal joint).
At present there is no complete Australopithecus foot. The composite shown here (Fig 1) is a best attempt, but it is a chimaera.
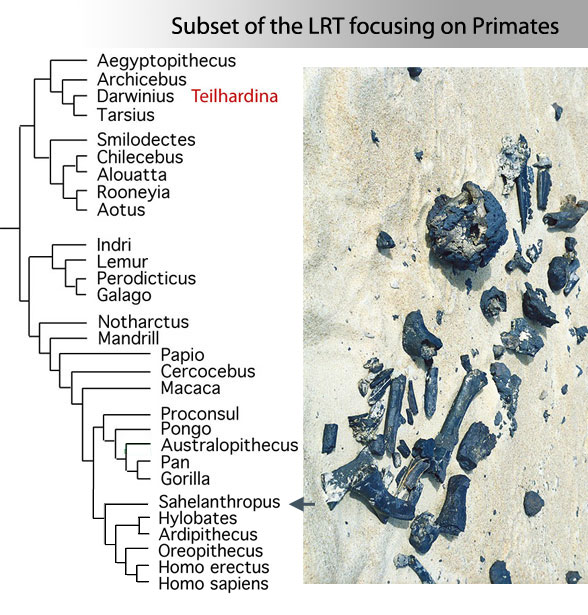
Figure 2. Subset of the LRT focusing on primates. Sahelanthropus in situ at right. A traditional primate, Microcebus, the mouse lemur, nests with bats in the LRT.
Not one of these traits, nor a dozen of these traits will answer which of these taxa are closer to humans. To do that every trait from nose to toe must be considered. That’s what the large reptile tree (LRT, 2082 taxa, subset Fig 2) does. Even so it produces only a hypothesis of interrelationships, subject to change with the addition of new taxa and score corrections. Presently the gibbon lineage leads to humans. The traditional human ancestor, Australopithecus, nests with apes.
References Brown P et al 2004. A new small-bodied hominin from the Late Pleistocene of Flores, Indonesia. Nature 431(7012): 1055–1061. Gierlinski GD et al (8 co-authors) 2017. Possible hominin footprints from the late Miocene (c. 5.7 Ma) of Crete? Proceedings of the Geologists’ Associatiion 128(5–6):697–710. Peters D 2000a. Description and Interpretation of Interphalangeal Lines in Tetrapods. Ichnos, 7: 11-41. Peters D 2010. In defence of parallel interphalangeal lines. Historical Biology iFirst article, 2010, 1–6 DOI: 10.1080/08912961003663500
Everyone knows microbats hunt with echolocation. They send out sound pulses from their larynx then listen for the faint echos from surrounding objects. In this way microbats create a ‘radar image’ of their surroundings. This is ideal for a nocturnal hunter. This is a trait convergent with tenrecs, then perfected in their descendants, the odontocetes (= toothed whales).
Today we’ll take a look at the ectotympanics of several bats and their ancestor looking for trends (Figs 1, 2).
Figure 1. Graphic depiction of several bats, ancestors and relatives to scale. Yellow highlights the ectotympanic, the bone shell that frames and contains the eardrum and middle ear bones. Full scale @72dpi screen resolution.
Like many clades of vertebrates bats had a phylogenetically miniaturized genesis (Fig. 1). The Mexican free-tailed bat (Tadarida, Figs. 1, 2) is the smallest bat tested in the Large Reptile Tree (LRT, 2082 taxa. Like its ancestor, Microcebus (Figs 1, 2), the tail is long and free, a primitive trait. Both also have a relatively large ectotympanic (eardrum frame). Note: as the skull phylogenetically gets smaller, the ectotympanic becomes relatively larger. Thus, the eardrum frame has a smaller variation in size than the skull it is attached to among these related taxa. Size matters.
Figure 2. Several bat skulls displayed here to the same snout-occiput length showing proportional changes to the skull in phylogenetic order. Ectotympanic is shown in yellow. µCT scan for Myotis from Nojiri et al. 2021.
Given the extra large ectotympanic in the LRT bat ancestor, the mouse lemur, Microcebus, do you wonder if it is capable of echolocation? Hending , Holderied and McCabe 2017 recorded their sounds. They wrote, “the vocal behavior of nocturnal primates has been poorly studied and there are few studies of their vocal repertoires. We compiled a vocal repertoire for the Endangered Sambirano mouse lemur, Microcebus sambiranensis.”
Their charts show Microcebus whistled in the 20kHz range. The human range of hearing is 20Hz to 20kHz.
The authors wrote, “Our equipment in this study limited our sampling rate to a maximum of 96 kHz, meaning that we lost some of the upper harmonics in our recordings. In future studies, we will use equipment capable of a sampling rate double the highest target frequency for mouse lemurs, i.e., a sampling rate of 200 kHz, which will ensure the upper harmonics are not cut from our recordings.”
Not sure how their charts scored into the 30–40kHz range given their stated limitations.
Figure 3. The Mexican free-tailed bat, Tadarida, is the most primitive extant bat tested in the LRT.
PS Tadarida was rescored for having a prefrontal and postfrontal. Scoring changes moved it toward the primitive bats in the LRT (Fig 4). Prior LRT subset cladograms will not reflect this change, but the LRT itself will. Always consult the LRT for the latest tree topology. It changes in subtle ways with every new taxon and as scores are corrected. The overall structure remains secure after eleven years of adding taxa and correcting scores.
Figure 4. Subset of the LRT focusing on bats.
References Hending D, Holderied M and McCabe G 2017. The use of Vocalizations of the Sambirano Mouse Lemur (Microcebus sambiranensis) in an Acoustic Survey of Habitat Preference. International Journal of Primatology 38(4):732–750. Nojiri T et al. (13 co-authors) 2021. Embryonic evidence uncovers convergent origins of laryngeal echolocation in bats. Current Biology 31(2):1353–1365.e3
Readers already know you cannot and should not trust everything you read, even if you read it here.
Science is all about testing and replicating. That goes for both data and results.Academic papers publish data, so readers can test the data. How do readers test data? One way (not the only way) is by adding taxa to cladograms, like the large reptile tree (LRT), the large pterosaur tree (LPT) and the therapsid skull tree (TST). Better yet, build your own cladogram from data you’ve studied, so you won’t have to borrow or trust someone else’s cladogram. It is always better to take ‘trust’, ‘belief’ and ‘reputation’ out of science. Stick with verifiable data.
For paleo authors: nobody likes to be pulled over, but a recent sampling of paleo papers (see below) have been speeding through the scientific method, peer review and publication process. Some of the most prominent seem to have kept their eye out for publicity opportunities, while overlooking self-examination and cold hard analysis that would have made their discovery less than superlative. Think of all the article headlines you’ve seen lately with ‘largest’, ‘oldest’, ‘first’, ‘bizarre’ and ‘baffled’. If you can’t think of any, a few are provided below.
Grant money and academic prestige must be associated with making paleo headlines. Otherwise paleontologists wouldn’t be interested in publicity. That’s the way it was for dozens of decades until the Internet era.
Scientists should welcome challenges to their results. That’s why they publish results, to see if they are valid or not. That’s called peer review whether it occurs before or after publication. Replication turns their hypothesis into a theory. Refutation is also part of the process. Unfortunately, and all too often, invalid methods, hypotheses and clades (e.g. genomic testing, volant azhdarchids and Avemetatarsalia) can become widespread and popularized because textbook writers and tuition dependent professors attempt to suppress challengers and keep those funds coming in. If results are wrong or are based on omitting pertinent taxa, scientists should admit their mistakes, then lift their chins again and carry on. Reputations should not be ruined by mistakes. Name calling is inappropriate.
In a worst case scenario, peer review after publication forced the retraction of Xing et al. 2020, the introduction of Oculudentavis, the lepidosaur originally described as an archosaur.
Retraction is not good. Subsequent papers should simply present arguments against published results. If all mistakes similar to Xing et al. 2020 had to be retracted for the same reason, every paper that listed pterosaurs as archosaurs should be similarly retracted. That hasn’t happened and it ain’t gonna happen.
The solution: Authors, editors and referees should raise their standards so mistakes like the following are published less often.
Recently the publicity universe was inundated with news about: The ‘largest Jurassic pterosaur’. Turns out Dearc was actually the fourth largest Jurassic pterosaur, and not a new genus.
Jagielska N et al. (ten co-authors) 2022. A skeleton from the Middle Jurassic of Scotland illuminates an earlier origin of large pterosaurs. Current Biology 32:1–8.
A hummingbird-sized dinosaur captured in amber. Turns out Oculudentavis was a lepidosaur (and a late-surviving cosesaur in the LRT).
Xing L, O’Connor JK,; Schmitz L, Chiappe LM, McKellar RC, Yi Q and Li G 2020. Hummingbird-sized dinosaur from the Cretaceous period of Myanmar. Nature. 579 (7798): 245–249.wiki/Oculudentavis
Bizarre bat-wing bird(s). Turns out Yi and Ambopteryx actually had (respectively) broken and crushed antebrachia (= ulna, radius).
Xu X, Zheng X-T, Sullivan C, Wang X-L, Xing l, Wang Y, Zhang X-M, O’Connor JK, Zhang F-C and Pan Y-H 2015. A bizarre Jurassic maniraptoran theropod with preserved evidence of membranous wings. Nature (advance online publication) doi:10.1038/nature14423 Wang M, O’Connor JK, Xu X and Zhou Z 2019. A new Jurassic scansoriopterygid and the loss of membraneous wings in theropod dinosaurs. Nature 569:256–259.
Lagerpetid ancestors of pterosaurs. Turns out these ‘ancestors’ were cobbled together from incomplete specimens found around the world, one of which ran on two toes, none of which resembled pterosaurs, other than being bipedal. No pterosaurs were bipedal according to the same misinformed authors who omitted pertinent taxa.
Ezcurra MD et al. (17 co-authors) 2020. Enigmatic dinosaur precursors bridge the gap to the origin of Pterosauria. Nature (2020). https://doi.org/10.1038/s41586-020-3011-4.
Müller RT 2022. The closest evolutionary relatives of pterosaurs: what the morphospace occupation of different skeletal regions tell us about lagerpetids. The Anatomical Record. https://doi.org/10.1002/ar.24904
Other invalid hypotheses: The genesis of baleen whales from toothed whales and toothed whales from artiodactyls. The genomic nesting of bats with horses and rhinos. The genomic nesting of flamingos with grebes. The genomic nesting of ducks with chickens. The placement of placoderms basal to sharks and bony fish. The origin of turtles from placodonts, diapsids and/or eunotosaurs. The traditional clades ‘Pseudosuchia’, ‘Pterodactyloidea’ and ‘Orinithodira’. The list goes on…
Long time readers will, no doubt, have their own favorite examples of authors hitting the publicity accelerator before taking a moment of reflection in the months to seconds prior to hitting the [upload] button. If your manuscript headline includes a superlative, do yourself and the rest of us a favor. Have someone else proofread and test it for you.
According to Wikipedia, “It is currently accepted that Marrella(Fig 1) is a stem group arthropod. Despite its superficial similarity to the trilobites, it is no more closely related to this group than it is to any other arthropod.Marrellahas too many antennae, too few cephalic legs, and too few segments per leg to be a trilobite.”
Depends on how you count them. The most common animal in the Burgess Shale, Marella (Fig 1) is way too specialized to lack a simpler ancestor.
Figure 1. Marella in several views from Garcia-Bellido and Collins 2006. Colors added here. Compare to the typical trilobite in figure 2.
Let’s start with the basics. Here’s a typical Cambrian trilobite (Triarthrus, ventral view, Fig 2). Color coding makes comparison of the many appendages and segments easier.
Too many ‘antennae’? No. Same number. One yellow. One red. Too few cephalic legs? No. And that changes all the time (Fig 3). Too few segments per leg? One fewer = nit-picking. Legs are rarely known in trilobites.
Figure 2. Triarthrus the trilobite, ventral view. Colors added here. Amber = prey grabbing/grinding-gill-legs. Green, olive and cyan = gill-legs. Compare to Marella in figure 1. Note the red appendages are anterior to the oral opening as in Marella,
According to Wikipedia, “Walcott described Marrella informally as a “lace crab” and described it more formally as an odd trilobite. It was later reassigned to the now defunct class Trilobitoidea. In 1971, Whittington undertook a thorough redescription of the animal and, on the basis of its legs, gills and head appendages, concluded that it was neither a trilobite, nor a chelicerate, nor a crustacean.
Figure 3. Another oddly derived trillobite, Dicranurus. Note the Marella-like cepahlic horns by convergence.
Invertebrate paleontologists report trilobites are extinct. That’s not true. Trilobites are in the ancestry of crustaceans, spiders and insects. Once again, this is low-hanging fruit, easy to discover just by coloring the appendages (Figs 1, 2).
References Garcia-Bellido DC and Collins DH 2006. A new study of Marrella splendens (Arthropoda, Marrellomorpha) from the Middle Cambrian Burgess Shale, British Columbia, Canada. Canadian Journal of Earth Science 43:721–742. Walcott CD 1912. Cambrian Geology and Paleontology 2, No. 6. Middle Cambrian Branchiopoda, Malacostraca, Trilobita and Merostomata. Smithsonian Miscellaneous Collections, 57(6):145–229. Whittington HB 1971b. Redescription of Marrella splendens (Trilobitoidea) from the Burgess Shale, Middle Cambrian, British Columbia. Bulletin of the Geological Survey of Canada, 209:1–24.
Yesterday the traditional primate, Microcebus (Fig. 1 right), untraditionally nested just outside the Primates in the large reptile tree (LRT, 2082 taxa, subset Fig 3), at the base of a bat + tree shrew + dermopteran clade called Volitantia. Pteropus, the flying fox (Fig 1 left) and Onychonycteris (Fig. 2) are clade members.
Today tree shrews and dermopterans move out, shifting closer to the gnawing clade in the LRT (Fig 3). Neither have large canines, similar to members of the gnawing clade. This is distinct from mouse lemurs, bats and primates, so that phylogenetic shift makes sense and is supported by additional scores from snout to rump.
Figure 1. Left: Pteropus the flying fox fruit bat. Right: Microcebus the mouse lemur.
In the most recent basal placental subset of the LRT (Fig 3) Microcebus nests at the very base of the bat clade (amber). This clade is a sister to the primate clade. That may be why primitive Microcebus is traditionally considered the smallest known primate. Linneaus 1758 recognized the similarity of bats and primates. Others (e.g. Pettigrew et al. 1989) have proposed similar hypotheses. The LRT nests bats, primates, dermopterans, tree shrews, shrews, rodents and multituberculates close to one another, sharing an unknown last common ancestor similar to Microcebus likely in the Early Jurassic.
Figure 2. Onychonycteris is a basal bat with large eye sockets.
Basal bats are even smaller than mouse lemurs. So phylogenetic miniaturization helped create basal bats from Microcebus-like ancestors.
The omnivorous bat + primate clade is the sister clade to the increasingly herbivorous clade of gnawing taxa (Fig 3). These clades are sisters to the insectivorous to piscivorous clade of anagalids + odontocetes (blue). All these are derived from omnivorous pre-Carnvora (= basal placentals) in the LRT.
Figure 3. A large moniphyletic clade in the LRT that includes mice, men, bats and dolphins.
This latest edit of the LRT narrows the gap between the basalmost bats and their most proximal non-volant outgroup, the mouse lemur, Microcebus. That means, at present the mouse lemur provides the best model for how pre-bats evolved, lived, behaved, reproduced, sounded, hibernated, etc.
Figure 4. Left: foot of Microcebus, demonstrating flexible ankle as in primates and bats. Right: foot of Green River bat. Even though the hallux (red) is not divergent, metatarsals 1 and 5 remain the widest. This and the large unguals on the bat are reversals going back to Nasua, the coatimundi, a by-product of neotony and phylogenetic miniaturization.
See how many of these Microcebus traits remind you of bats: The eyes are large, typical of nocturnal mammals. The mouse lemur forages alone, but sleeps in groups, sharing tree holes during the day. Mostly insectivorous, plus small reptiles, frogs, plants, leaves, fruits and flowers. Habitat: dense tropical forests. Twin babies are typical and are carried in the mother’s mouth after birth, prior to weaning. Offspring can reproduce after one year with a lifespan of ten years. Pteropus and Microcebus have similar eyes, faces, nostrils (Fig 1).
Genetics Both the mouse lemur (Microcebus) and the unrelated aye-aye (Daubentonia) live in Madagascar along with many dozen lemur species. That’s why their genes, affected by endemic deep time viruses, align with those of lemurs.
The LRT is always under scrutiny especially so as new taxa are added. Scores from years ago get a review every so often. It’s an ongoing process. The present changes seem to make sense. Let me know if you notice any problems, omissions, etc.
This appears to be a novel hypothesis of interrelationships. If not, please provide a prior citation so I can promote it here.
References Linnaeus C 1758. Systema naturæ per regna tria naturæ, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Tomus I. Editio decima, reformata. Miller JF 1777. Cimelia Physica p.25 Pettigrew JD, Jamieson BGM, Robson SK, Hall LS, McAnally KI and Cooper HM 1989. Phylogenetic Relations Between Microbats, Megabats and Primates (Mammalia: Chiroptera and Primates. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 325 (1229): 489-559. David Attenboroughmeets a mouse lemur video.
Today’s blogpost admittedly starts slow and seemingly off subject, but it rounds the bases and comes back to paleo home plate by the end.
Ever hear the phrase? “Ahh… that’s the kind of thing you do when you’re 14 or 15…“
Yaron Brook on Lex Fridman podcast 138
Yaron Brook makes the point: “those who say that… have lost something. They’ve lost their youthful idealism.”
Brook continues, “What makes youthfulness meaningful? Sometime in the teen years, there’s something that happens in the consciousness, we almost awaken anew. We suddenly discover we can think for ourselves. We suddenly discover that not everything our parents and teachers tell us is true. We suddenly discover that this tool, our minds, is suddenly available to us to discover the world and to discover truth. And it is a time of idealism. And it’s a time of I want to know about the world. I want to go out there. I don’t believe my parents. I don’t believe my teachers.
And this is healthy. This is fantastic. And I want to go out there and experiment. And that gets us into trouble. We do stupid things when we’re teenagers. Why? Because we’re experimenting. It’s the experiential part of it. We want to go and experience life. But we’re learning. It’s part of the learning process and we become risk takers because we want experience. There’s a risk with something we need to learn because we need to learn where the boundaries are.”
“One of the damages that helicopter parents do is they try to prevent us from taking those risks so we don’t learn about the world and we don’t learn about where the boundaries are.”
I think I understand Dr. Naish in a new way now. He was trying to be a helicopter parent. He was telling his readers that something bad was out there on the Internet. That something bad had happened to me as I built the large reptile tree (= LRT, then several hundred taxa, now 2080). Naish was trying to warn his readers that I was discovering and documenting novel relationships outside of Benton’s Vertebrate Paleontology textbook.
Here’s the point: Naish was taught from Benton’s textbook. Naish teaches from Benton’s textbook. He knows Benton personally. Naish was trying to protect his future ability to attract paleo students and make a living teaching from Benton’s textbook. That’s understandable for a dad and husband. Naish was raising a family and that always hits the core.
Admittedly, ten years ago at age 58, I was acting like an experimenting 14-year-old by starting to build the LRT. At least it seems so now, after hearing what Brook has to say.
Brook continues, “So the teenage years are years of wonder. They’re depressing when you’re in them. But they are exciting periods of discovery and people get excited about ideas. Good ideas. Bad ideas. All kinds of ideas.”
“And then what happens?”
“We settle. We compromise. Think about what academia does for so many of us. We go into academia and we’re excited. We’re going to learn stuff. And we’re going to discover things. And then they stick you into some sub-field, examining some minutia that’s insignificant and unimportant and to get published you have to be conventional. You have to do what everybody else does. And then there’s the tenure process of seven years where they put you through this torture to write papers that fit into a certain mold. By the time you’re done you’re in your mid 30s and you’ve done nothing. You’ve discovered nothing. You’re all in this minutia and this stuff and it’s destructive.”
“Holding on to that knowledge and confidence is hard. And when people do away with that they become part of the system. And they inflict the same pain on the next guy that they suffered because that’s part of how it works.”
Inflicting ‘the same pain on the next guy.’ Now who would do that?
“It’s in every field. It’s asking the big questions.“
‘The big questions‘ are answered by adding taxa to the LRT.
“It’s staying curious. It’s staying excited and accepting failure. You’re not going to get it the first time. You’re not going to get the whole thing. And sometimes you have to do the minutia work. And I’m not here to say nobody should specialize. You have to do that. There has to be a way to keep it all integrated.”
Integration is the central concept behind the LRT: a wider gamut of taxa, from fish to birds, are tested together in the LRT than have ever been tested together before.
“We don’t live in a culture that integrates. We don’t have a holistic view of these things. You’re not going to make the big leaps forward without a holistic view.”
PhDs may not like it, they may not cite it, but the LRT has made big leaps forward with nothing more than a holistic view. When PhDs employed the same taxon lists, they got the same results. In hind sight those several times now appear to have been convergent accidents. Now that more PhDs know better, they don’t employ the same taxon list as the LRT as often in recent years (see the Timeline of Pterosaur Origin Studies blogpost).
Remember Bennett 2012 restricted his pterosaur origin study to archosauromorphs. That was twelve years after several phylogenetic analyses removed pterosaurs from archosaurormorphs and five years after pterosaurs were moved into Lepidosauria along with Huehuecuetzpall (Peters 2007). That’s not an exception. That’s typical. There is little interest in testing results that are published outside the current paradigms in Benton’s textbook.
In vertebrate paleontology the LRT is that long sought holistic view. The LRT integrates millions of data points and minutia. It continues to be an ongoing experiment built from errors and omissions revealed by corrections and additions. Every time another taxon is added to the LRT it becomes more holistic, more integrated, better able to document the gradual accumulation of derived traits between sister taxa. As expected, older posts are not as accurate nor as complete as more recent posts.
The LRT is widely disliked even though it resolves enigmas and documents a gradual accumulation of traits across all included taxa. At present no other published cladograms out there include such a wide gamut of chordate taxa.
And that is why the LRT needs competition. It needs a dozen similar studies to test LRT results and expose errors here, there and everywhere. Since the LRT is disliked, let’s find somewhere out there a trait-based study with a similar gamut that is liked.
Build your own LRT with errors and omissions, corrections and additions. You’ll find it the most powerful tool in your toolbox. When you do this, let us know your results and share your .nex files. Let’s figure this evolution thing out together. Don’t settle. Don’t compromise. Keep that teen spirit of experimentation and that distrust of authority. This is science, not religion. You’re supposed to question everything.
You’ll know you’re on the wrong track when your critics provide hard evidence and additional taxa that falsify your claims. Acting like patient teachers, the good critics will patiently guide you to tenable solutions.
You’ll know you’re on the right track 1. when your critics discuss your freshman mistakes, include surrealistic graphics from others (rather than your own tracings) and conclude that nothing you offer has any value. 2. when your critics ask you to add characters (a never-ending, never-enough request). 3. when your critics say things like “Face palm!” (don’t call them snowflakes) 4. When your critics get sarcastic, hyperbolic, over emotional (60% of the comments section) or resort to name-calling and labeling. When those things happen, don’t fire back. Calmly and simply remind your critics that they are scientists or wannabe scientists. Science is for everyone. Not just for PhDs.
After all, professors and wannabe professors, like all adults, are just big kids who pay taxes, drive cars and also worry what others think about them. Sometimes workers succumb to peer group pressure and think they have to defend the castle. When they do, gently remind them that they are not acting like the little open-to-experimentation scientists they think they are. If you know such a scientist who has settled and compromised, remind them of their youthful idealism. Encourage them to build a competing LRT or do something that is even more integrated, holistic and parsimonious.
References Bennett SC 2012. The phylogenetic position of the Pterosauria within the Archosauromorpha re-examined. Historical Biology. iFirst article, 2012, 1–19. Peters D 2007. The origin and radiation of the Pterosauria. In D. Hone ed. Flugsaurier. The Wellnhofer pterosaur meeting, 2007, Munich, Germany. p. 27. Yaron Brook on the Lex Fridman Podcast
Pittman et al. 2022 support the old and invalid pterosaur quad launch on water hypothesis (Fig 1) first proposed by co-author Habib 2008, who imagined a catapult mechanism presented by him online here and shown in figure 2 (left).
Figure 1. GIF animation from the American Museum of Natural History showing how their Pteranodon managed to hop off the surface of the water until suddenly able to flap and fly. Physics? That’s not pertinent in this animation.
Unfortunately that hypothesis was undercut by imagined pterosaur morphology (Fig 2 left). Habib imagined planting that big wing finger tendon on the ground to get an elastic snap/catapult. Habib imagined the three anterior/medial free fingers had migrated to the former posterior/lateral surface of metacarpal 4 axially rotated to the dorsal side in pterosaurs (keeping finger 4 flexing in the plane of the wing). Habib imagined shortening the three free fingers until they lost contact with the substrate (Fig. 2 left).
Figure 2. Left: The so-called catapult mechanism in pterosaurs. Right. The actual design of pterosaur (in this case Anhanguera/Santandactylus) fingers. Click to enlarge.
In the real world pterosaur prints demonstrate finger 4 never contacts the substrate. The three free fingers never sit on the axially rotated dorsal side of metacarpal 4. They make contact with the substrate. The three small fingers always extend beyond the big wing joint as shown in figure 2 right. Every pterosaur enthusiast knows this, but apparently no one else has told Habib he has made those mistakes. Instead they embrace Habib and his mistakes. He’s a co-author on Pittman et al. 2022.
The Pittman et al. title, “Quadrupedal water launch capability demonstrated in small Late Jurassic pterosaurs” turns out to be either a matter of misdirection or a hoax. When you get inside this paper there is no demonstration. There is no reconstruction of the pterosaur doing what the authors hypothesize it doing (Fig 1). There is no reconstruction at all. Additionally, there are no alternate scenarios presented and weighed one against the other. There is no science here. There is only invalid speculation by Pittman et al. 2022 based on invalid speculation by Hone and Henderson 2014.
Pittman et al. report, “Most living water birds use a dynamic running launch.” Is that true? No. Even the relatively large pelican (video above) first flaps free of the water, then starts building up forward motion with more flaps and feeble, relatively ineffectual paddles with its webbed feet.
Figure 3. At left Pteranodon traced from bones floating in two configurations, each with two water levels based on hollow bones throughout, including those pontoons called wings. At right completely imagined Pteranodon from Hone and Henderson 2013 (2014), which Pittman et al. 2022 took for data. c
Pittman et al. base their hypothesis on an earlier paper, Hone and Henderson’s 2014, whose inability to understand how pterosaurs, the least dense of all vertebrates, were able to float on water. That foible is illustrated by them above (Fig. 3 right). This is especially egregious when Hone and Hennderson can’t float their Pteranodon, which carries its own long, hollow metacarpal pontoons. Hone and Henderson freehand their imagined generic Pteranodon model rather than following the more precise instructions provided by a complete Pteranodon skeleton (Fig 3 left). Purportedly acting like scientists, even an amateur can tell Hone and Henderson didn’t do their due diligence. They muddled through and got published. This paper should have died an obscure death, but today…
Pittman et al. were published in Nature based on that 8-year-old mistake. Citing Hone and Henderson, Pittman et al. report, “A purportedly low floating position“. Beyond all reason, Pittman et al. are admitting the data they are basing their report on is “purported” (= little more than flimsy hearsay). That is not a good starting point in science. They don’t bother to test that old hypothesis either. Instead Pittman et al. borrowed an inaccurate, largely imagined conclusion to make their preconceived point (= quad launch from water). Well, that’s they way it is nowadays. Actually that’s the way paleo has been for the last two decades. Evidence doesn’t seem to matter.
Whatever happened to testing? To observing nature? If workers are going to borrow, that’s where they should be spending their time, outside observing, not locked up in a study imagining things that never were and never could be.
Getting back to a pterosaur water launch: How do ducks do it? How do pelicans do it? How do kingfishers do it? Built from a convergent but similar bauplan to birds, that’s how pterosaurs did it. They flapped from a floating or underwater position developing both thrust and lift to free themselves from the water, like extant ducks, pelicans and kingfishers. Pterosaurs didn’t stand on their folded wings, patting, spanking or slapping the water (or ground) to get airborne (contra figure 1). That way lies a dangerous crash and face-plant.
Sorry these myths keep getting recycled by PhDs. They must be under some sort of peer group pressure to do so. The rest of us can only do our best to bring reason and understanding back to pterosaurs (Fig 3 left), especially when all of us venture beyond the bones into behaviors.
References Habib MB 2008. Comparative evidence for quadrupedal launch in pterosaurs. Zitteliana B28, 159–166. Habib M and Cunningham, J 2010. Capacity for water launch in Anhanguera and Quetzalcoatlus. Acta Geosci. Sin. 31, 24–25 (2010). Habib M. http://pterosaur.net/anatomy.php Hone DWE and Henderson DM 2014. The posture of floating pterosaurs: Ecological implications for inhabiting marine and freshwater habitats. Palaeogeography, Palaeoclimatology, Palaeoecology 394:89–98. Pittman M, Kaye TG, Campos HB and Habib MB 2022. Quadrupedal water launch capability demonstrated in small Late Jurassic pterosaurs. Nature Scientific Reports 12:6540. https://www.nature.com/articles/s41598-022-10507-2
Following long on the heels of decades of earlier dino-documentaries (list below) here’s the latest and perhaps greatest attempt at taking viewers back to the Cretaceous. The CGI is first rate (based on the previews = trailers).
Link to YouTube trailer for Prehistoric Planet. Click to play.
Figure 1. Cast and crew of Prehistoric Planet includes these lumnaries and a mass of CGI artists.
According to writer, Rachel Funnell: “Set to air on Apple TV+ in May, Prehistoric Planet’s features, designs, and information were all consulted by scientists including palaeozoologist Darren Naish, and science illustrator and paleoartist Gabriel Ugueto. This means for the first time feathery baby T.rexes, patterned Pachyrhines, and even sauropod throat sacs are on display”
Every CGI dinosaur looks and moves as if everyone on board did their homework. They are all excellent! Of course, and typically unfortunately, the pterosaurs did not get the same attention (Fig 2). They seem to have been done in a more imaginative, freehand manner with little to no regard for the skeleton inside. Here a skeleton of the most complete azhdarchid, Quetzalcoatlus lawsoni, is shown alongside and only the outstretched wings overlaid for comparison.
Figure 2. Quetzalcoatlus sunning itself, distorted somewhat, according to the more gracile skeleton added here standing beside it. The producers so much wanted this giant to fly, not realizing it became a giant only after becoming flightless, as in so many giant, but first flightless birds.
Some journalists are saying this is supposed to be a CGI Hatzegopteryx on the beach. Since relatively little is known of Hatzegopteryx (e.g. cervicals, partial humerus, partial dentary), that provides wide latitude for restoring the rest of the animal with imagination. If this is Hatzegopteryx, let’s look at some of what little is known of this Romanian pterosaur.
Figure 3. Added April 21, 2022 with photo images of Quetzalcoatlus in comparison to Hatzegopteryx.
Here (Fig. 3) is the Hatzegopteryx humerus to scale with Q northropi and Q lawsoni. Missing parts for Hatzegopteryx are restored based on Q northropi from Andres and Langston 2021. The diameter of the shaft at the break indicates the Hatzegopteryx humerus was overall larger than Q northropi, likely creating a larger pterosaur. Now, look at the deltopectoral crest of Hatzegopteryx. It is relatively reduced. That means the pectoral flapping muscles were smaller in the largest pterosaur. That means Hatzegopteryx was probably non-volant considering the potentially larger overall size of Hatzegopteryx and the inability of the smaller azhdarchids to fly based on their overall proportions. These include the oversized head on the end of a relatively stiff neck, the reduced wings, reduced wing phalanges and reduced sternal complex (Fig 1) on Quetzalcoatlus. Panic flapping was still an option that could have added thrust to speedy runs across the terrain.
According to Wikipedia, “The documentary follows dinosaurs recreated with computer-generated imagery living around the globe in the Late Cretaceous period, 66 million years ago.[2] It is the first major dinosaur-focused documentary series produced by the BBC since Planet Dinosaur in 2011, and the third overall (the first was Walking with Dinosaurs in 1999). The series uses up-to-date palaeontological research in order to depict its animals of the Cretaceous with scientific rigour; for example, some species featured in the previews are covered in feathers, such as a juvenile Tyrannosaurus rex.”
Link to YouTube trailer for Prehistoric Planet. Click to play.