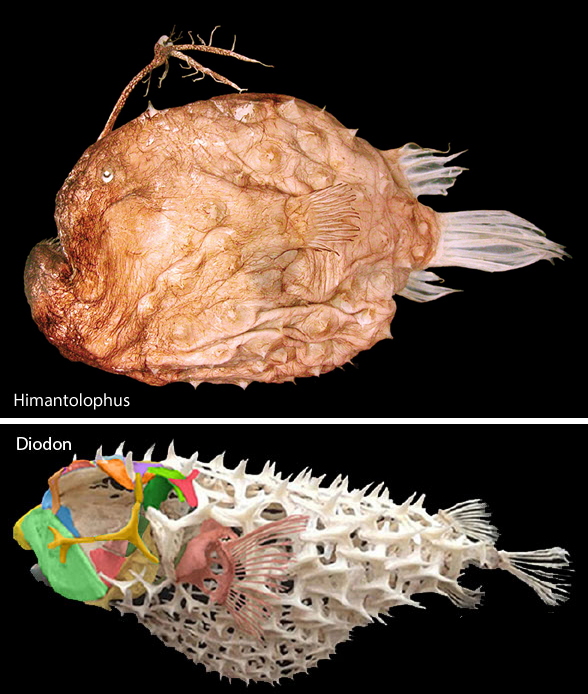
Short one today. More housekeeping in the fish subsection of the large reptile tree (LRT, 2297 taxa) is driving taxa together that have not been nested together before. Today the football fish,Himantolophus, nests with the porcupine fish, Diodon (Fig 1) in the LRT.
Himantolophus is a traditional anglerfish. In the LRT it is now convergent with anglerfish, which are closer to scorpionfish and perch.
Seems obvious now, especially noting that Himantolophus also lacks pelvic fins and has dermal spines all over its body. This hypothesis of interrelationships could change tomorrow. And that’s OK.
Himantolophus groenlandicus (Reinhardt JCH 183; 60cm in length) is a deep sea ‘angler’. Strongly resembling the frogfish Antennarius, Himantolophus lacks any trace of a pelvis or pelvic fin. In the LRT it is more closely related to Diodon the porcupine fish (Fig 1). The skin is covered with large bony plates, each with a medial spine. The teeth are depressible. This single genus has 18 species.
BTW, the LRT has not been updated for the past week and cannot be updated for awhile considering the current wave of changes, like this one (Fig 1), coming into the fish subset of the LRT.
Thank you for your readership, yes, even you certain readers, who hate DGS, the LRT and every time I post here. I realize this is frustrating for you.
Solution: Build your own LRT. Then you’ll have evidence to counter hypotheses made here and elsewhere. Become a player. It’s more fun than sitting in the stands.
Long time readers have been following this 12-year phylogenetic experiment in real time, including mistakes, discoveries, corrections and new hypotheses of interrelationships. There has been only one reason to build the LRT: it has never been done before, not with this wide gamut of 2297 taxa (Chordates). This analysis needed to be done because taxon exclusion is the number one problem in vertebrate paleontology.
There is no need to cite hypotheses presented here. Just add pertinent taxa to your own more focused studies.
References Reinhardt JCH 1837. Ichthyologiske bidrag til den Grönlandske fauna. Kgl. Danske Vidensk. Selsk. Natur. Math. Afhandl., 7: 83-196, pls. 1-8.
Today we return to a controversy that began a few days ago when the vertebral series of Perucetus (Figs 1–4) was introduced to the world. Many videos and articles followed this announcement.
Science writer Flora Graham wrote an editorial for Nature on Perucetus Here’s the subhead with a modern tie-in. “Perucetus colossus, an ancient whale that lived 38 million years ago, is a new contender for the heaviest animal ever. Plus, four key questions on the new wave of anti-obesity drugs.”
I’ve never seen Nature add a commercial to their subheads before. Funny that they connected “anti-obesity drugs” to the “heaviest animal ever.”
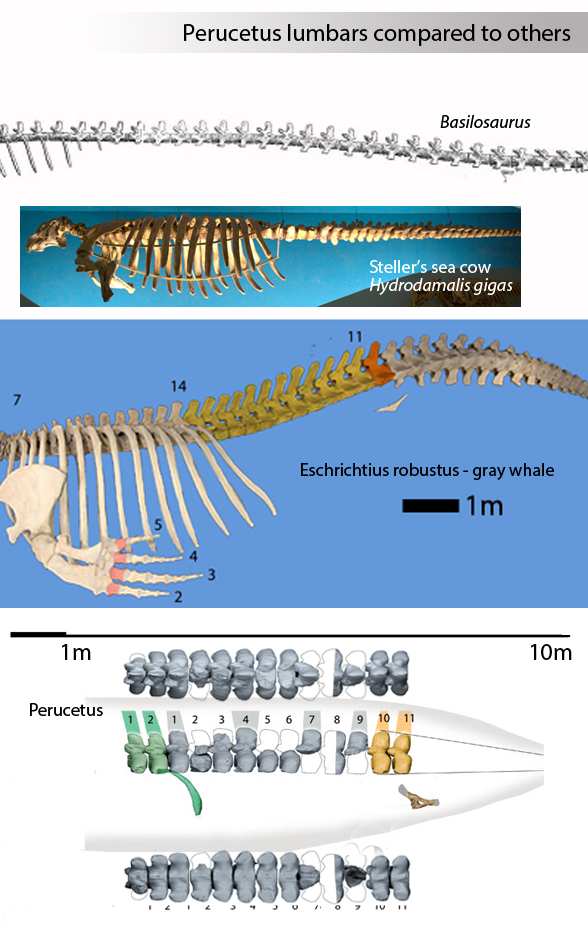
Figure 1. Comparing an average Balaenoptera and the only Perucetus lumbar vertebrae. The extant mysticete has a much larger centrum cross-section. The extinct taxon has much more robust transverse processes, adding to the weight of the bone, likely for bottom-feeding in the clam beds it was found in. Note the blue whale vertebrae is shown anterior to the right, reversed from the Perucetus vertebrae, anterior to left.
Getting back to science. Perucetus and its authors (Bianucci et al 2023) have more than a few followers. Among them is science writer, Flora Graham, in an editorial for Nature. Her headline, “Giant ancient whale could be heavier than blue whale” is ambiguous, but juicy. Graham didn’t say ‘heavier than the largest ever blue whale’, which measured 110ft or 33.5m. The average size for the blue whale is 80 to 90 feet long, shorter for males.
So there’s lots of wiggle room here to boost that Perucetus headline.
Figure 2. How to estimate the size of something if adult human males are your only yardstick. I’m tall so my ulna is 29cm long. The average male has a shorter ulna: 27.4cm..
Facts and estimates Weight of largest ever blue whale: 418,878 pounds or 209 tons. What Graham wrote for Nature:“Most blue whales weigh around 100–150 tonnes; the researchers’ best guess is that Percetus colossus weighed around 180 tonnes.” Wikipedia reports for Perucetus, “weight ranging from 85–340 t (84–335 long tons; 94–375 short tons),”
So there’s lots of wiggle room here, too, especially for a set of Perucetus lumbars (Fig 3) that are not larger than those of an average blue whale (Fig 4). Bianucci et al recognized these estimates were wide-ranging, to their credit.
Figure 3. Comparing three different scale bars vs the Perucetus vertebrae. The longest possible measurement of the known bones of Perucetus based on published scale bars indicates a length of 4.6 meters for the set. So published scale bars were within the realm of correct. Measuring with human yardsticks (Fig 2) gives a shorter value for the lumbar series. Note the neural spine orientation changes in this series, distinct from other whales (Fig 4).
Environomental writer, Emma Marris, also writing for Nature had this to say: “might have been heavier than a blue whale, even if it was not as long.” “The whale’s bones were so enlarged that they look swollen, almost like inflated balloons.” “The bones lacked the porous, spongy structure of typical bones.”
The last two are true. The first is a big maybe not based on current evidence (Fig 4).
Figure 4. Perucetus compared to Basilosaurus, Balaenoptera and Cynthiacetus to scale. As shown in figure 3, the last two vertebrae tapered toward the tail. So reconstructions that show a longer line of lumbars, as in basiolosaurids, do not appear to fit the bones. Short neural spines are found in Desmostylus (Fig 7). Perucetus has a transitional length of neural spines. Perucetus had shorter thoracic ribs than Balaenoptera.
Bone density is related to buoyancy. Densely-boned animals tends to sink. Whales of all types, despite their less dense vertebrae, generally sink when dead, providing islands of nutrition on the sea floor for years thereafter. By contrast, right whales tend to float, at least at first, which made them the ‘right’ whales to harpoon back in the day. Hippos are able to walk on river floors at least partly due to denser than typical limb bones. The marine hippos, the desmostylians, had a presumed similar lifestyle grazing the shallow sea floor, even though they had whale-like bone density in their vertebrae (Hayashi et al. 2013).
If Perucetus grazed the sea floor aided by heavy bone density, and, at best, vestigial hind limbs, we might assume it lived and fed in waters no deeper than its nostril height – which means rather shallow waters. Perhaps other factors enable it to float over deeper waters.
Interesting parameters.“Although manatees are excellent swimmers, the deepest an individual has been seen diving is 10 m (33 ft.). They normally feed no deeper than about 3 m (10 ft.) below the surface of the water.
Why did Bianucci et al focus on Late Eocene Cynthiacetus peruvianus (Fig 4) for their Perucetus Bauplan? Both are from Peru and both have similar morphologies if one simply expands each vertebra in Cynthiacetus to match Perucetus. Why did Bianucci et al not consider a desmostylian or mysticete connection? Probably because all the published literature ignores desmostylians as candidate ancestors for mysticetes. Most of the published literature considers archaeocetes basal to all extant whales.
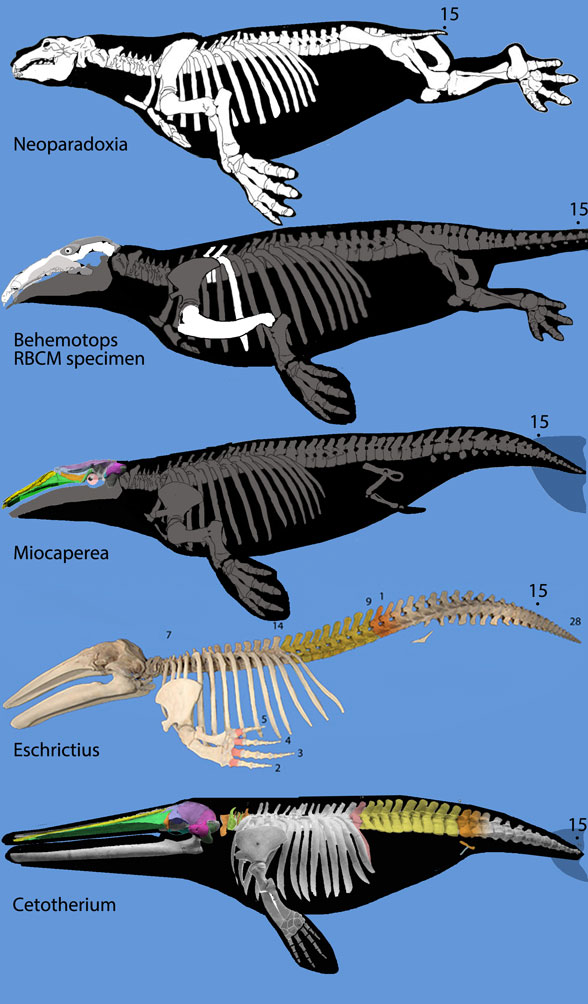
Figure 5. Rorqual evolution from desmostylians, Neoparadoxia, the RBCM specimen of Behemotops, Miocaperea, Eschrichtius and Cetotherium, not to scale. Note the tightly packed lumbars of Cetoterhium, much like Perucetus. Dark gray bones are hypothetical based on phylogenetic bracketing.
Short pre- and post-zygopophyses on Perucetus (Fig 3) would appear to indicate a short range of motion. The brevity of the transverse processes is a second indicator. The brevity of the neural spines is a third indicator. Given these parameters, perhaps the tail of Perucetus was not the massive organ of propulsion seen in basilosaurids and extant whales that have taller, longer, narrower, lighter vertebral processes (Fig 4). Given these parameters the authors considered Perucetus to be a slow moving marine mammal, like a manatee. Along the same lines, let’s not omit the possibility of a very large desmostylian transitional to its own lineage of mysticetes (Fig 5) based on that massive lumbar series and tiny pelvis (Fig 7).
The Barnucci et al SuppData indicated greater flexibility in the lumbar region: 26º dorsal, 60º indicated ventral – but 160º demonstrated ventral, and 17º lateral flexion.
Distinct from other whales of all types, the Perucetus neural spines did not all lean the same way (Fig 3). They diverged in the front and back. In other mysticetes, Ziphius and Orcinus, all the neural spines lean posteriorly. In terrestrial mammals, Delphinus and deep-diving Physeter, the sperm whale, the neural spine angles converge over the anterior caudals.
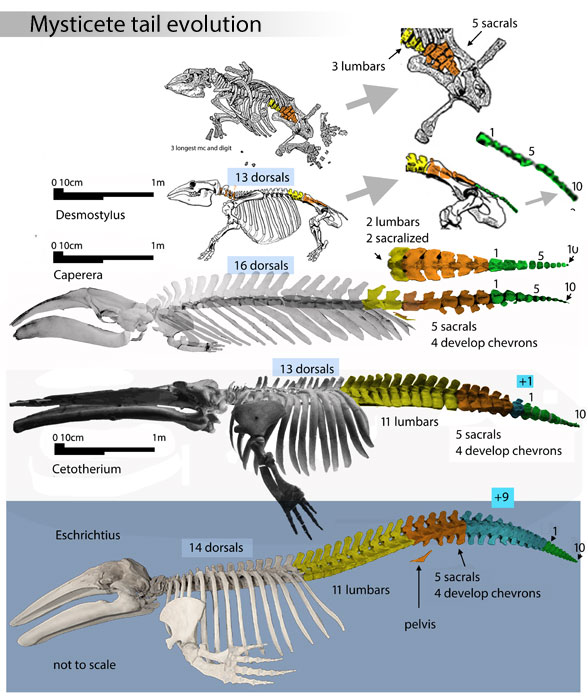
Figure 6. The origin of a robust tail in the ancestor of mysticetes (baleen whales). Note the paddle-like ribs on Caperea and Cetotherium, like those found with Perucetus (figure 4). Note the short, low transverse processes on the long lumbar region of Cetotherium and Eschrichtius AND Perucetus – but not the same in Caperea, which has a short lumbar region and more elongate centra.
Miocaperea (Fig 5 middle) is a Miocene mysticete or pre-mysticete from Peru. Too bad it is only known from a skull. We can only speculate on the post-cranial anatomy. Too bad Eocene Perucetus is only known from vertebrae. We can only speculate on the cranial anatomy. Both are considered bottom-feeders in shallow waters based on the phylogenetic proximity of Miocapera to desmostylians and the extreme density of the Perucetus vertebrae. Let’s keep open the possibility that these two taxa might be related to one another.
Figure 7. Desmostylus skeleton compared to Perucetus lumbars and pelvis, not to scale. The short neural spines are similar to Perucetus, but the high transverse processes are more primitive in Desmostylus. The pelvis shapes are similar, but the sizes are not. Note the acetabulum contacts the anterior margin in both taxa. The LRT nests desmostylians basal to mysticetes.
Here’s an interesting fact with phylogenetic significance. “Eight months after conception, at the height of the wet season, female hippos give birth to one calf at a time, either on land or underwater. Afterwards, mothers leave the herd for a short period of time to bond with their calves underwater. After a few weeks, the calves finally exit the water to feed on grass.”
Since mysticetes give birth only underwater, we can assume transitional taxa, like desmostylians, moved increasingly to giving birth underwater.
Miocaperea pulchra (Bisconti 2012; late Miocene, 7–8 Ma; 40cm skull length) was considered a Miocene ancestor of Caperea. Here it nests between Behemotops (Fig 5) and Eschrichtius, the gray whale (Fig 5). The post-crania in Miocaperea is not known, so it could have had legs or flippers.
YouTube video in Spanish. Turn on Closed Caption (cc) and translate to English or your favorite language in YouTube settings.
The above 1:11 hour YouTube video is informative, if not a little rambling, joking and opinionated. To his credit, the narrator emphasizes all numbers are estimates. To his credit the narrator realizes the environment must have been rich in food for Perucetus to grow so large.
References Bianucci G et al (15 co-authors) 2023. A heavyweight early whale pushes the boundaries of vertebrate morphology. Nature. doi:10.1038/s41586-023-06381-1 Hayashi S et al (8 co-authors) 2013.Bone Inner Structure Suggests Increasing Aquatic Adaptations in Desmostylia (Mammalia, Afrotheria). PlosOne https://doi.org/10.1371/journal.pone.0059146 Graham F 2023. Daily briefing: Giant ancient whale could be heavier than blue whale. Nature Daily Briefing Online here. Marris E 2023. Could this ancient whale be the heaviest animal ever? Nature News Online here. Peters D unpublished. The triple origin of whales. ResearchGate.net georgiasfossils.com/12c-cynthiacetus-revised
Just another experiment trying to figure things out. Here the Early Triassic crocodylomorph, Dyoplax, was traced again using DGS methods. Then missing and disturbed parts were restored (Fig 1) based on sister taxa in the large reptile tree (LRT, 2297 taxa, subset Fig 2). Those sisters are (at present) Late Jurassic Alligatorellus (Fig 1) and extant Gavialis (Fig 1).
Figure 1. Alligatorellus, Dyoplax and Gavialis compared. Missing and disturbed parts of Dyoplax are restored here.
Dyoplaxarenaceus (Fraas 1867, Lucas, Wild and Hunt 1998; Maisch, Matzke and Rathgeber 2013) is a unique Late Triassic crocodylomorph, one of the three original crocodylomorphs in the 19th century (along with two aetosaurs). Dyoplax nests in the LRT as a basal gavialid and close to Jurassic sea crocodiles.
Alligatorellus beaumonti (Gervais 1871; Late Jurassic) was found in Solnhofen marine limestones. It is a small, short-snouted primitive carnivore, closely related to Late Triassic Dyoplax and the gharial, Gavialis. It has juvenile proportions, so could represent a neotonous adult.
Figure 3. Subset of the LRT focusing on Crocodylomorphain 2023. The new nestings represent just the latest of a changing hypothesis of interrelationships.
A paper on Gavialis phylogeny (Gold, Brochu and Norell 2014) reported, “Some areas of the tree remain controversial, particularly near the base of the crocodylian evolutionary tree. Among these is the phylogenetic position of the Indian gharial, Gavialis gangeticus, whose phylogenetic placement changes dramatically depending on which types of data are analyzed.”
The authors used phylogenetic analysis and genomic analysis. They reported, “The braincase data alone produced a clade that included crocodylids and Gavialis,whereas the Eustachian data resulted in Gavialis being considered a basally divergent lineage.” The authors concluded, “In this study, when the morphometric data were added to discrete morphological and molecular characters, they each result in the identical topology in which Gavialis and Tomistoma combined are closest relatives within Crocodylidae.”
By contrast, in the LRT (Fig 3) using trait analysis Tomistoma (the false gharial) nests basal to Alligator and Crocodylus, apart from Gavialis (the gharial).
Data is not always complete pristine and incontrovertible. Even so, use the data you have and keep looking for improvements, even if they upset / modify your current hypothesis. Run the numbers in your own LRT to find out for yourself.
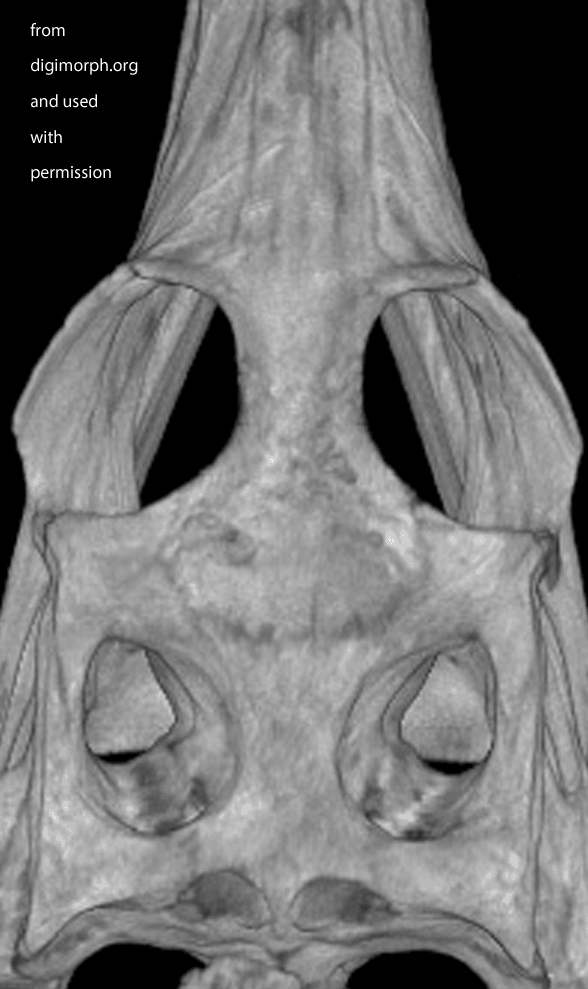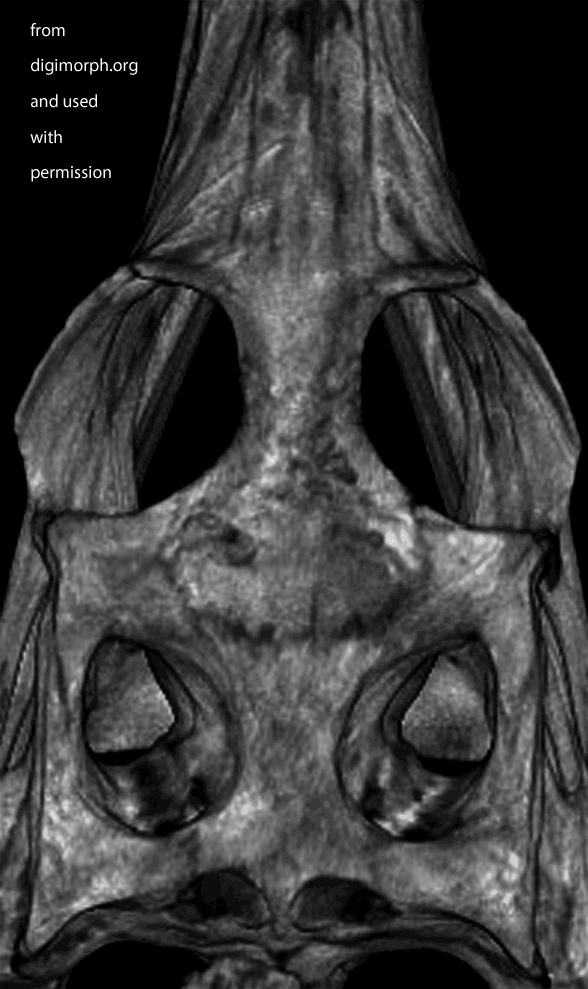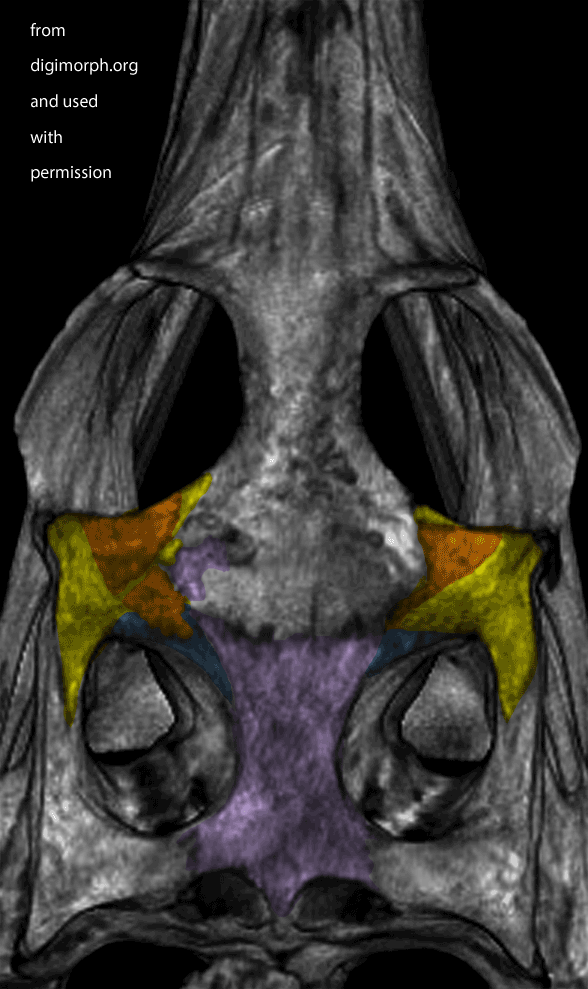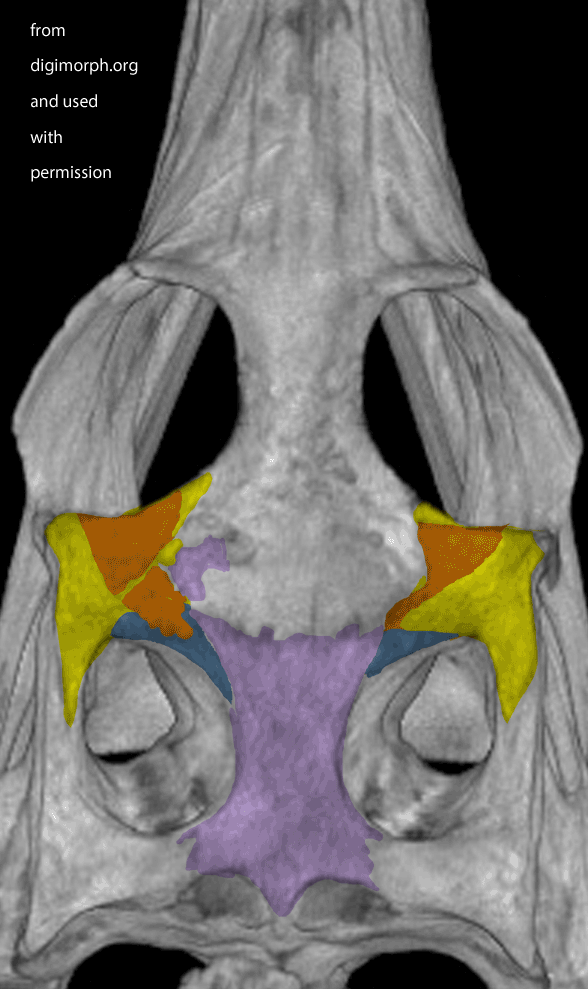
PS. Croc expert and paleontologist Christopher Brochu questioned the colored division of the traditional croc postorbital into postorbital (yellow) and postfrontal (orange) regions in figure 1 above. He wrote, “Crocodylians don’t have postfrontals at any point in development. They’re not fused to the postorbital – they’re absent entirely. And even if the two bones were fused (which they’re not, since one of the two bones never appears), I have no idea how you would distinguish them on an actual skull. I would recommend re-coloring the postorbital as a single bone, because that’s what it is.”
A closer look at the bones should be educational. My reply and request follow.
Figure X. Dorsal view of a Gavialis skull from Digimorph.org. Frame 2 boosts contrast. Frame 3 adds colors to indicate where I see a postfrontal (orange) on top of the postorbital (yellow). The left side appears to have suffered a bit of damage (missing part of the frontal to expose the underlying parietal in lavender).
Thanks, Chris, I appreciate your extensive experience with crocodilians. To your point, the vast majority of derived crocodylomorphs have obliterated the suture between the postorbital and postfrontal. To call the remaining bone a postorbital is appropriate because the postfrontal appears to phylogenetically shrink relative to the postorbital.
The LRT nests Gavialis apart from crocs + gators, so Gavialis might not follow crocodilian rules. Brochu 1997 also split Gavialis apart from other crocodilians. That paper did not include Alligatorellus and Dyoplax (Fig 1). The LRT nests Gavialis with these hatchling-sized adult taxa that appear to have a suture between the postorbital and postfrontal. As always, it is hard to tell.
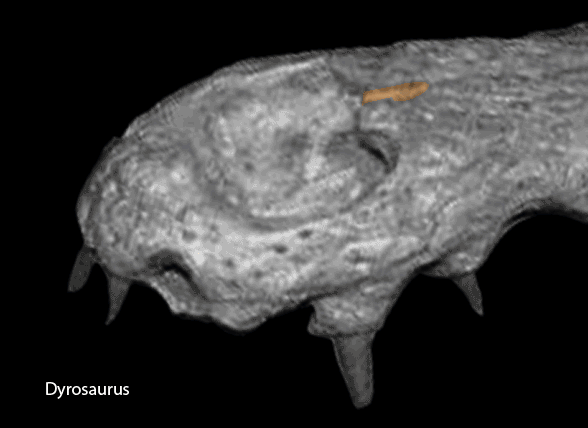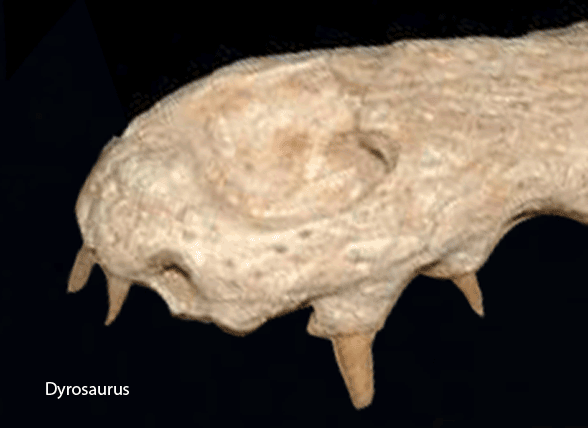
A few crocodylomorphs appear to keep the suture between the postfrontal and postorbital. Dyrosaurus (Fig y) related to Gavialis in the LRT, Fig 2) appears to be one.
Figure Y. Dyrosaurus appears to keep a separation between the postfrontal and postorbital in this adult specimen. It is related to Gavialis in the LRT.
Can you provide evidence of a postorbital-postfrontal complete fusion (obliterated suture) in a gharial that matches the seamless condition in crocs and gators?
While waiting for your evidence, I posted an image from Digimorph.org of a Gavialis skull in dorsal view (Fig x). Frame 2 boosts the contrast. Frame 3 colorizes what little remains of the postfrontal largely atop the postorbital. The bones are fused. Faint outlines appear to remain. There appears to be some damage on the left.
Rather than posting a comment on this image, it would be better if you repeated this tracing exercise and send the results for posting here,. That way we will have two players on the field, rather than one player and one umpire.
I sought images of a Gavialis hatchling skull, but none were found.
PPS – a day later Chris Brochu replied, “If you can demonstrate that any of the points I’ve made is inaccurate, please let me know.”
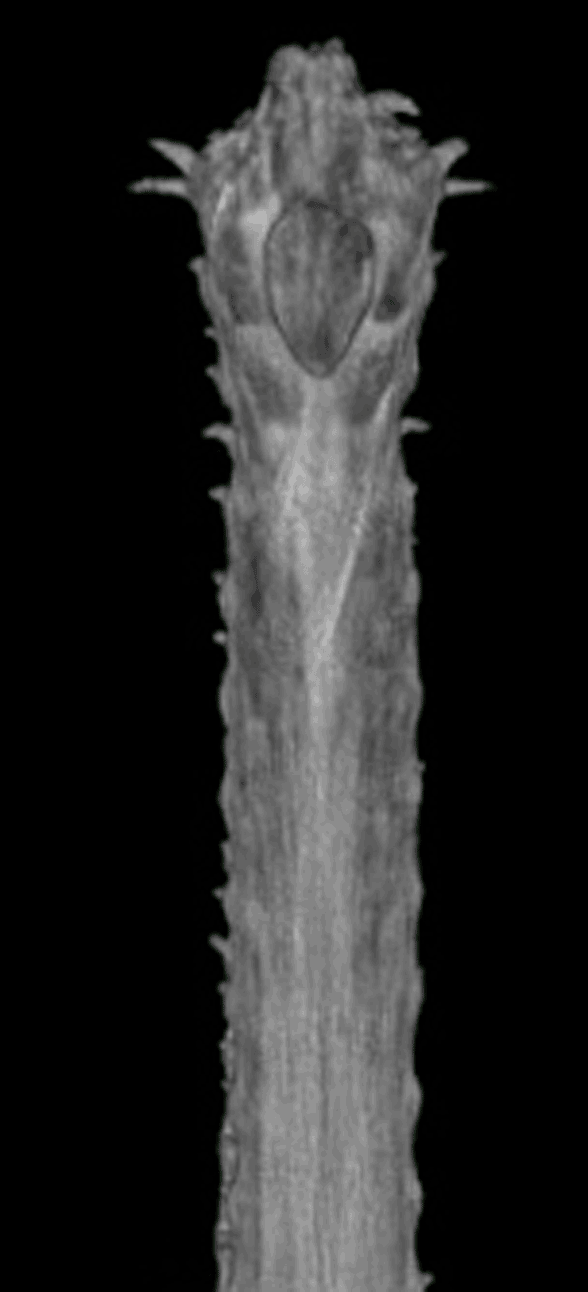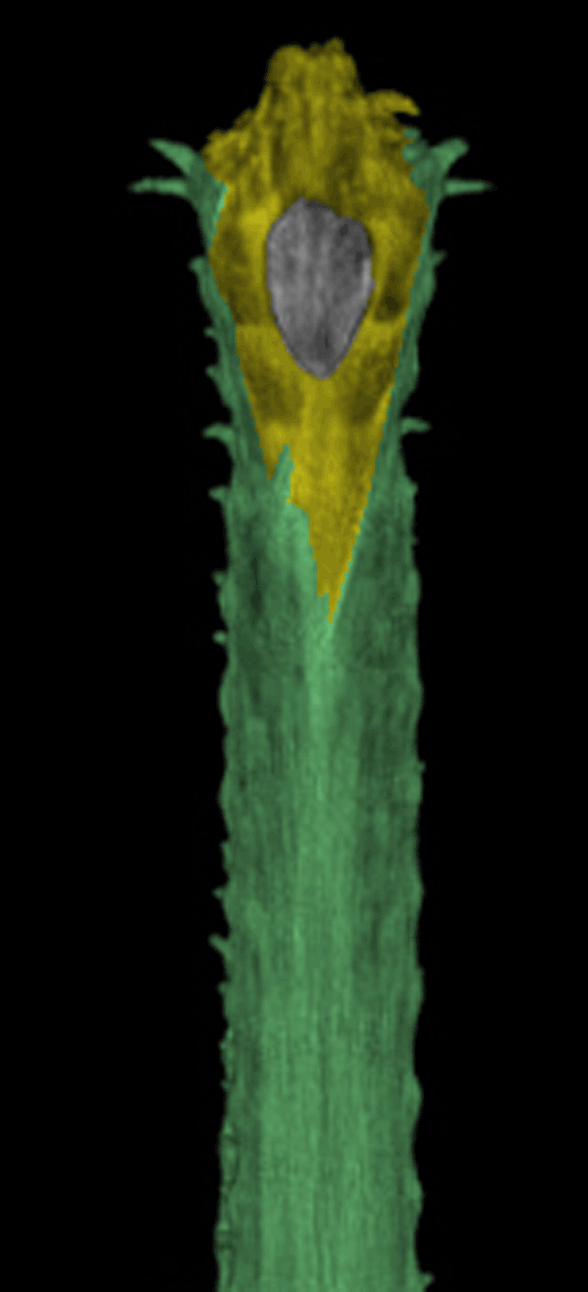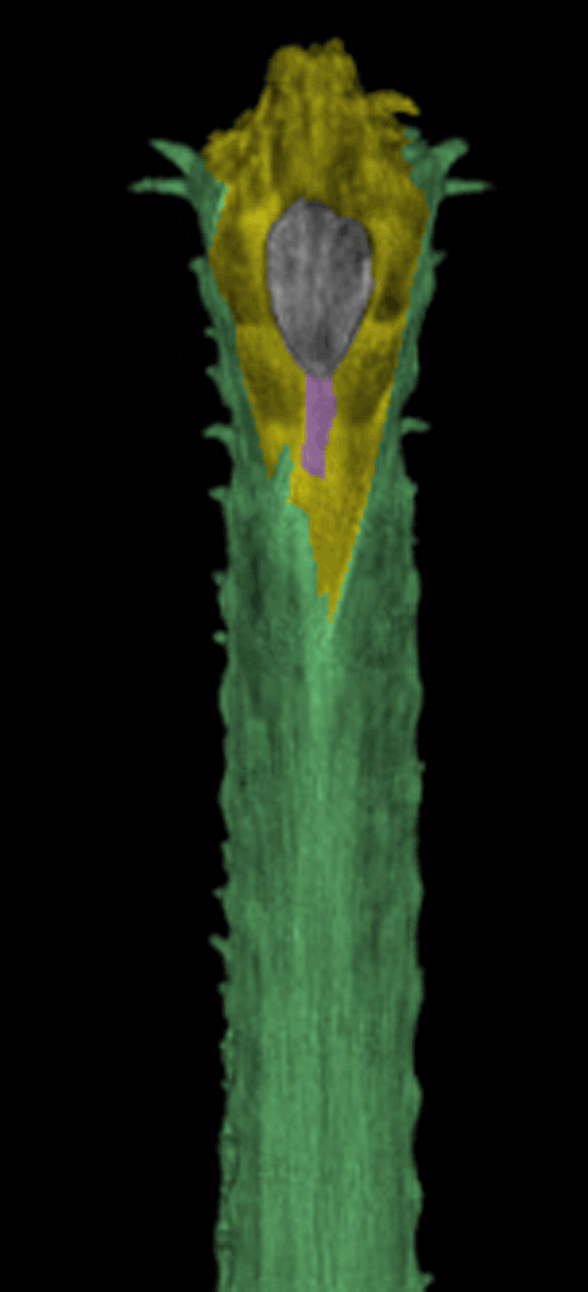
Well here’s the snout of Gavialis. This is the demonstration Brochu asked for. Frame 1 shows the Digimorph.org image alone. Frame 2 shows the snout with the premaxillae (yellow) blocking the nasals from contacting the naris, which is not seen in most tetrapods, including other crocodylomorphs, but is seen in certain extinct crocodylomorphs. Frame 3 shows the nasals (pink), if present, contacting the naris. If the nasals do not contact the naris, at least readers can see the reason why this appeared to be so from this image. See other related taxa shown below ((Fig YY, ZZ) for relevant homologies.
Figure XX. Dorsal view of Gavialis snout. Frame 2 shows the premaxillae blocking nasal contact. Frame 3 shows nasal (pink) contact, as in figures YY and ZZ. This is a younger specimen than the one shown below in figure x.
Figure YY. Snout diagram of Isisfordia showing the nasals contacting the naris.
Figure ZZ. Snout of Dyrosaurus showing the nasals contacting the naris in the manner of Gavialis.
At some point a judgement can be rendered. Are postfrontals present or not (Fig X)? Does the nasal contact the the naris or not (Fig XX)? Sometimes the answer is clear (Figs YY and ZZ). Sometimes the answer is not so clear (Fig XX).
This question is a microcosm of the DGS method. DGS can demonstrate observations to readers. On the other hand, when a scientist reprorts, “Having seen the specimen you’re colorizing… I can assure you that your sutural reconstructions are inaccurate” does not demonstrate observations to readers. It must be believed. No evidence or demonstration is necessary. Statements like this are appeals to authority. They should not be questioned, even with evidence.
For many people, appeals to authority are attractive. That’s why we still have bat-wing birds. Some readers will latch on to Brochu’s appeal to authority in order to blackwash the entire LRT and the DGS method. That is regrettable, but that happens.
I prefer to present evidence, however tenuous (Fig 1). I agree, the evidence for the nasals contacting the naris in Gavialis is tenuous, at present, depending on shadows and light and homologies in closely related taxa. A µCT scan should be able to determine whether of not the nasals beneath the maxillae and premaxilla still contact the naris – or not. It might vary among individuals. So there’s a solution to this argument. Don’t make this a tempest in a teapot. We can figure this out, or let it drop.
It’s not life-or-death. It’s an idea.
FIgure x. Added more than a week later following a citation to this figure, which purports to show the truncation of the nasals in Gavialis. Note the nasals extend to the naris in the other crocodilians. Note also the tiny medial bone (pink arrow) in the top illlustration of the Gavialis snout in the exact place occupied by the nasals in other crocs. By this evidence the middle portion of the Gavialis nasal has been lost, leaving only a small anterior portion (extent unknown below the overlapping premaxilla) and a larger posterior portion, now separated from the anterior. Excellent use of DGS, by the way. Glad to see other workers adopting this technique for demonstration and teaching.
References Fraas O 1867.Dyoplax arenaceus, ein neuer Stuttgarter Keuper-Saurier. Jh. Verein vaterländ. Naturk. Württemberg 23:108-112; Stuttgart. Gold MEL, Brochu CA and Norell MA 2014. An Expanded Combined Evidence Approach to the Gavialis Problem Using Geometric Morphometric Data from Crocodylian Braincases and Eustachian Systems. PLoS ONE 9(9): e105793. doi:10.1371/journal.pone.0105793. Lucas SG, Wild R, Hunt AP 1998.Dyoplax O. Fraas, a Triassic sphenosuchian from Germany. Stuttgarter Beiträge zur Naturkunde, B. 263: 1–13. Maisch MW, Matzke AT and Rathgeber T 2013. Re-evaluation of the enigmatic archosaur Dyoplax arenaceus O. Fraas, 1867 from the Schilfsandstein (Stuttgart Formation, lower Carnian, Upper Triassic) of Stuttgart, Germany. Neues Jahrbuch für Geologie und Paläontologie – Abhandlungen. 267 (3): 353–362.
From the abstract “We provide direct dietary evidence that Jeholornis consumed leaves likely from the magnoliid angiosperm clade, and these results lend further support for early ecological connections among the earliest birds and angiosperms. Morphometric reanalysis of the lower jaw of Jeholornis further supports a generalized morphology shared with other herbivorous birds, including an extant avian folivore, the hoatzin.”
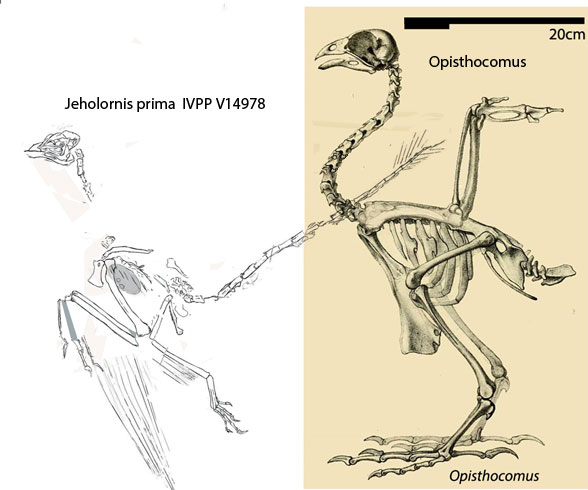
Indeed, the skull of Ophisthocormus, the hoatzin (Figs 1, 2), does greatly resemble (by convergence) that of the new Cretaceous plant-eater, Jeholornis (Figs 1, 2).
Figure 1. Jeholornis and Opisthocomus both ate plants by convergence.
The question is is the new Jeholornis prima conspecific with the holotype (Fig 3)? The holotype has teeth, which are lacking in the new specimen. A distinct skull shape is also notable.
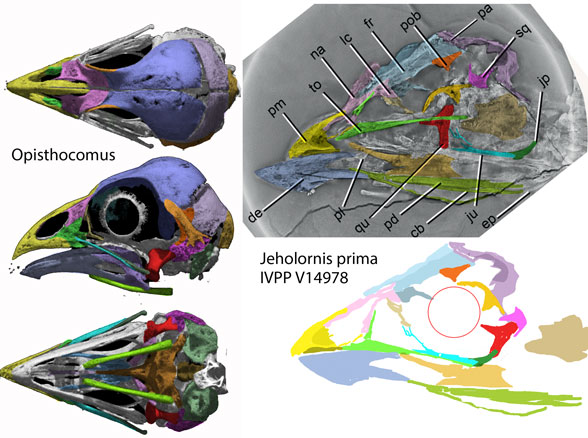
Figure 2. Opisthocomus skull in 3 views compared to the skull of Jeholornis in situ and reconstructed using DGS methods.
Figure 3. Jeholornis prima holotype compared to the new plant-eating specimen. Are they conspecific? Apparently no, but close enough to nest together in the LRT.
When added to the large reptile tree (LRT, 2299 taxa), the three Jeholornis specimens nest together. Their differences are not so great as to separate them = moving them closer to other taxa.
References Yan Wu, Yong Ge, Han Hu, Thomas A. Stidham, Zhiheng Li, Alida M. Bailleul and Zhonghe Zhou, 2023. Intra-gastric phytoliths provide evidence for folivory in basal avialans of the Early Cretaceous Jehol Biota by 28 July 2023, Nature Communications. DOI: 10.1038/s41467-023-40311-z
Publicity from SciTechDaily.com “New findings from the analysis of a 120-million-year-old fossil skeleton of the extinct bird Jeholornis, unearthed from northeastern China, present the earliest known evidence of leaf-eating birds, marking the earliest known evolution of arboreal plant-eating among birds.
“The pheasant-sized Jeholornis, a member of the second most primitive lineage of known birds, has teeth and a long bony tail like its predatory, feathered dinosaur relatives. However, microscopic analysis of the fossilized residues in the stomach of this juvenile, arboreal (tree-living) bird demonstrates that Jeholornis was not a predator. It had eaten tree leaves from a group of flowering plants called magnoliids that includes the living magnolia, cinnamon, and avocado trees.
“After comparison with over 4,000 kinds of modern phytoliths, we can see that most of the identifiable fossil phytoliths from the stomach come from the leaves of magnoliids,” said Dr. Wu Yan from IVPP, first author of the study.
“To further support their hypothesis of leaf-eating in this early bird, the paleontologists also compared the lower jaw of this bird to living birds with a wide range of diets. Coauthor Dr. Hu Han from Oxford University said, “A more detailed statistical analysis of the three-dimensional shape of the lower jaw of Jeholornis shows similarities to the shapes of living birds that eat mostly plants including the living leaf specialist, the hoatzin from tropical forests in South America.”
From the Fang et al abstract: “Modern baleen whales are unique as large-sized filter feeders, but their roles were replicated much earlier by diverse marine reptiles of the Mesozoic.
Replicated? Which taxa? If you can’t come up with a short list, readers will consider this sentence fluff = pushing a narrative lacking evidence or a list to check. Provide the evidence. Provide the list of diverse marine reptile filter feeders.
“Here, we investigate convergence in skull morphology between modern baleen whales and one of the earliest marine reptiles, the basal ichthyosauromorph Hupehsuchus nanchangensis, from the Early Triassic, a time of rapid recovery of life following profound mass extinction.
“Two new specimens reveal the skull morphology especially in dorsal view.”
“The snout of Hupehsuchus is highly convergent with modern baleen whales, as shown in a morphometric analysis including 130 modern aquatic amniotes.”
Notable by their absence: any taxa from the clade Mesosauria, which actually have baleen-like procumbent needle-teeth and phylogenetically precede hupehsuchids in the large reptile tree (LRT, 2297 taxa).
Build your own LRT to find this out for yourself.
“Convergences in the snout include the unfused upper jaw, specialized intermediate space in the divided premaxilla and grooves around the labial margin.’
Maybe not (See Fig 1).
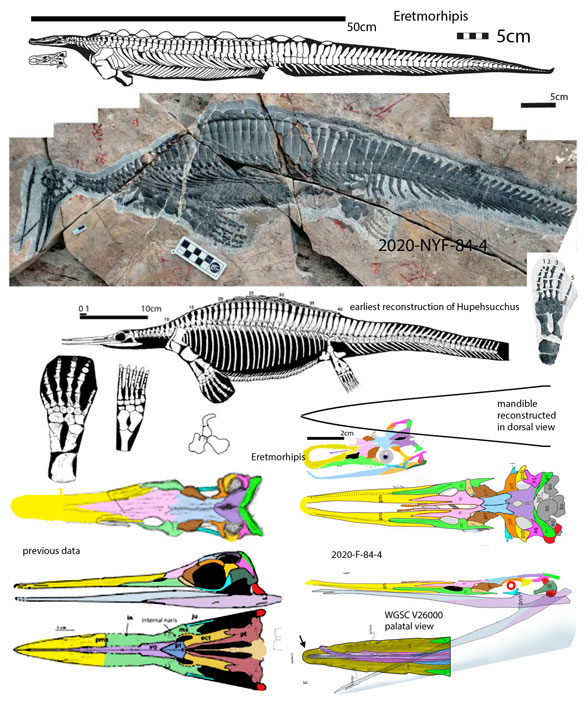
Figure 1. Huphesuchus, the 2020-F-84-4 specimen and Eretmorohipis compared. Note the medial fenestra and laterally oriented dorsal ribs in Eretmorhipis and the 2020-F-84-4 specimen. Look at the rostrum of the 2020-F-84-4 speicmen. Is it more like a kitchen strainer? Or more like a tweezers?
Unfortunately, when the new 2020-F-84-4 specimen was added to the LRT, it did not nest with Hupehsuchus (Fig 1), but closer to Eretmorhips(Fig 1).
Unfortunately the unfused upper jaw and divided premaxilla of 2020-F-84-4 is not shared with related taxa. This condition appears to be due to taphonomic damage at the tip. Undamaged specimens (Fig 1) retain that tip that bridges both sides of the premaxilla.
Unfortunately the ‘specialized intermediate space’ is shared with Eretmorhipis (Fig 1), not Hupehsuchus. Another related taxon, Eohupehsuchus, has an even narrower rostrum lacking any dorsomedial opening.
From the Fang et al abstract: “Hupehsuchus had enlarged its buccal cavity to enable efficient filter feeding and probably used soft tissues like baleen to expel the water from the oral cavity.”
The buccal cavity (= mouth) or the 2020-F-84-4 specimen is actually relatively smaller compared to its body than the holotype of Hupehsuchus (Fig 1). However, the mouth is indeed larger on the publicity illustration (Fig 2).
The buccal cavity of 2020-F-84-4 can be described as small, flat and pointed. Is there room for baleen-like filtering structures in the extremely shallow rostrum of the 2020-F-84-4 specimen? Maybe not. Are those needle-thin mandibles capable of bowing out like mysticete and pelican jaws? Maybe not. Usually if a taxon is gulping a lot of water, those jaws are really big. The jaws on this and related hupehsuchids are more like flat to pointed tweezers. They are particularly gracile on 2020-F-84-4 (Fig 1).
Figure 2. Two images of the same specimen. One for science. One for publicity. If you see any differences or exaggerations, so does everyone else. If you want your specimen to have a larger mouth rimmed with baleen and more robust limbs, just let the illustrator know. Is this good science? You decide.
The authors mention studying one platypus, which has a similar flat toothless, baleen-less buccal cavity, a similar overall morphology, a similar size and thus more likely a similar lifestyle. No further mention was made of that platypus in the text and its similarity to hupehsuchids in general. We looked at this topic a few years ago here.
From the Fang et al abstract: “Coordinated with the rigid trunk and pachyostotic ribs suggests low speeds of aquatic locomotion, Hupehsuchus probably employed continuous ram filter feeding as in extant bowhead and right whales.
Maybe not. Their figure 2 compares Hupehsuchus to a rorqual, a Minke whale, not a right or bowhead whale. If you say ‘right whale’ you should show ‘right whale’. If you show a Minke whale, you should say a MInke whale. And show them to scale
The Early Triassic palaeoenvironment of a restrictive lagoon with low productivity drove Hupehsuchus to feed on zooplankton, which facilitated ecosystem recovery in the Nanzhang-Yuan’an Fauna at the beginning of the Mesozoic.”
When a changing niche drives a taxon to a particular modification, you’re going to have to show some ancestors and some phylogenetic relatives. As mentioned earlier, Early Permian pre-mesosaurs were ancestral to hupehsuchids + ichthyosaurs in the LRT. So there’s a starting point worth examining, but overlooked by the authors.
Take a look at Wumengosaurus (Fig 3) for a hupehsuchid relative.
Another reason to run an analysis: Make sure your new taxon is indeed the genus and species you call it in your headline. This time a mistake was made.
Figure 3. From 2018. Basal Ichthyosauria, including Wumengosaurus, Eohupehsuchus, Hupehsuchus and Thesaurus, the taxon closest to ichthyosaurs.
What does a platypus eat? “They scoop up insects and larvae, shellfish, and worms in their bill along with bits of gravel and mud from the bottom. All this material is stored in cheek pouches and, at the surface, mashed for consumption. Platypuses do not have teeth, so the bits of gravel help them to “chew” their meal.”
Sounds like what a hupesuchid might do, even without baleen.
References Fang et al (8 co-authors including testbook author MJ Benton) 2023. First filter feeding in the Early Triassic: cranial morphological convergence between Hupehsuchus and baleen whales. BMC Ecology and Evolution. https://doi.org/10.1186/s12862-023-02143-9
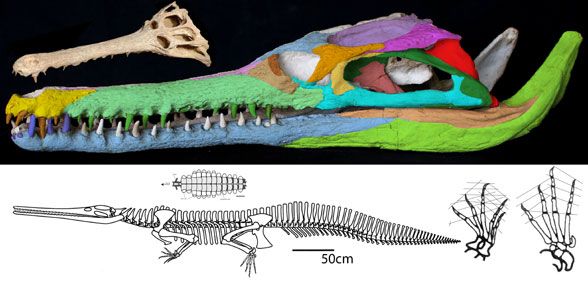
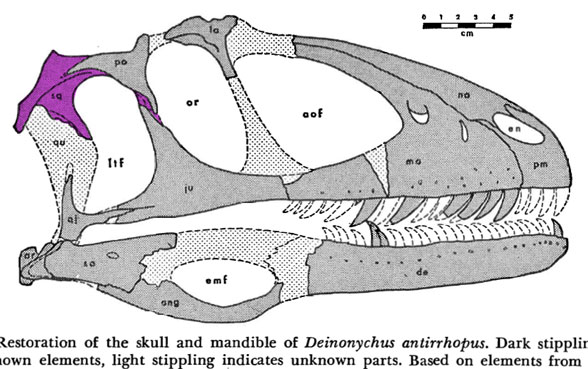
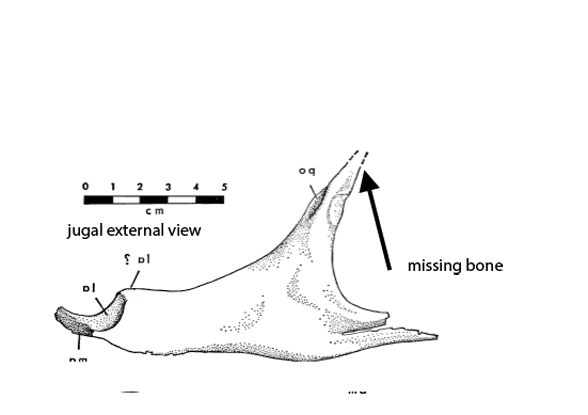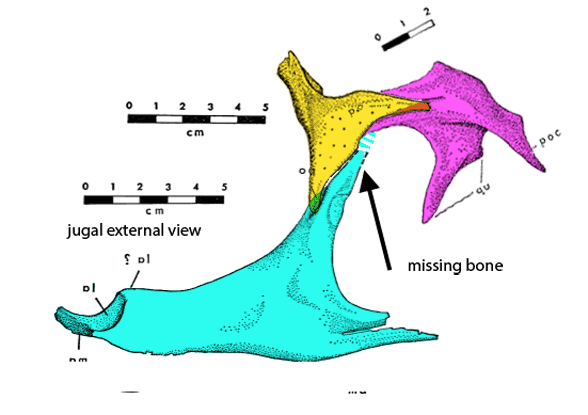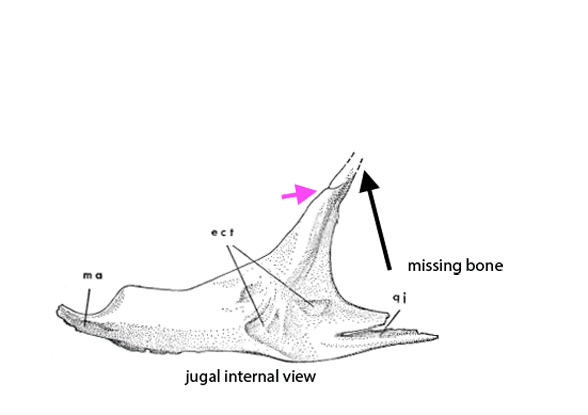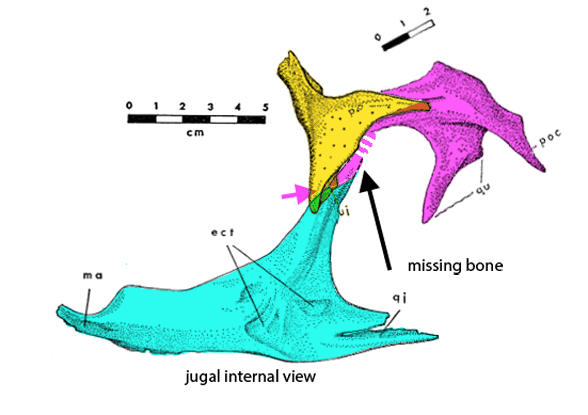
This is the dinosaur that started the dinosaur renaissance. Deinonychus antirrhopus(Ostrom 1969, Early Cretaceous, Figs 1–4) had the first ‘killer’ claw, a tendon stiffened tail and an Archaeopteryx-like manus (hand). These traits indicated a highly active, likely warm-blooded predator, more like an osprey (Pandion), than a Komodo dragon (Varanus).
Figure 1. The Harvard specimen of Deinonychus. This is not a complete specimen. Some preserved parts are detailed below.
Figure 2. Deinonychus skull. Colors added here. Note the long jugal-squamosal (cyan-magenta) contact.
Figure 2b. Figure from Ostrom 1970. Color added here to show the extent of the squamosal. Any differences between the original drawings and the sculptured skull are in the purview of the museum sculptors, who appear to have had access to the original materials.
Figure 2c. A preparator’s dilemma. Did the jugal contact the squamosal or not? The preparator made a choice to show contact (Fig 2). Ostrom’s 1970 drawing of the individual bones (assembled here) indicates vanishing bone at the key area. Here two possibilities for contact are shown. Of course, bone could have been absent here – but the model maker chose contact from data known to them. As readers know, corrections are part of the scientific process. This is correction #212,444 or thereabouts.
Distinct from tested relatives the orbit of Deinonychus (Fig 2) is deeper than wide and not wider than the post-orbital length of the skull. Note the long jugal-squamosal (cyan-magenta) contact is also distinct.
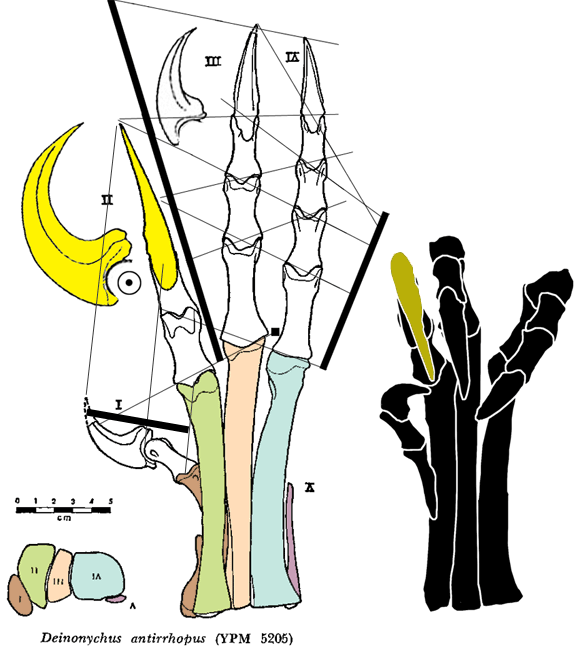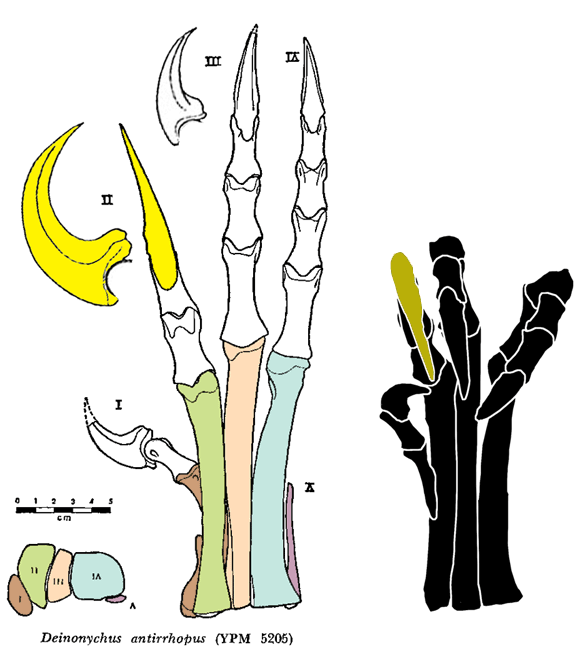
PILs (parallel interphalangeal lines) can be drawn through most, but not all of the phalangeal joints of the hand (Fig 3) and foot (Fig 4).
Figure 4. Deinonychus pes, extended and flexed in ventral view. Parallel Interphalangeal Lines (PILs) added here.
In the LRT Deinonychus is derived from Late Jurassic Ornitholestes and Late Jurassic Sciurumimus, neither of which had a ‘killer’ claw nor a chevron-tendon-stiffened tail.
References Ostrom JH 1970. Stratigraphy and paleontology of the Cloverly Formation (Lower Cretaceous) of the Bighorn Basin area, Wyoming and Montana”. Bulletin of the Peabody Museum of Natural History. 35: 1–234.
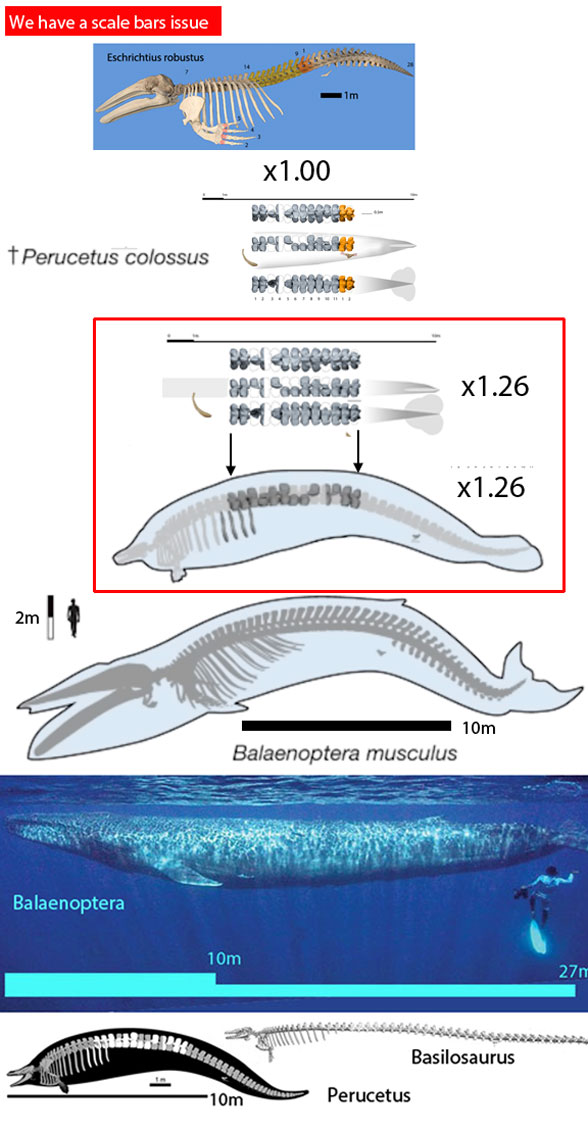
This Nature paper by Bianucci et al 2023 includes several superlatives (e.g. ‘heaviest animal on record‘) that do not pan out. Mismatching scale bars are the issue. One set of scale bars (on the published restoration, Fig 1) makes Perucetus about 26 percent larger (Fig 1) than the second set of scale bars on the bones alone (Figs 1, 2). Accident? Oversight? Or some other motivation?
Weight estimates range from 85 to 340 tons. That’s a wide range! A good scientist will always want to emphasize the high side – if you want headlines!
Turns out Perucetus may have been a robust extinct relative of the gray whale (genus; Eschrichtius robustus (Figs 1, 2). This is only a guess. Too few traits were preserved to add Perucetus to the LRT.
Figure 1. Perucetus presented two time to scale with 27m long (90 foot) blue whale and 20m long Basilosaurus. Is Perucetus the largest or heaviest animal ever? Not according to the scale bars. The larger version of the two Perucetus specimens is inflated by about 26 percent.
From the abstract: “The fossil record of cetaceans documents how terrestrial animals acquired extreme adaptations and transitioned to a fully aquatic lifestyle. In whales, this is associated with a substantial increase in maximum body size. Although an elongate body was acquired early in cetacean evolution, the maximum body mass of baleen whales reflects a recent diversification that culminated in the blue whale”.
In the large reptile tree(LRT, 2295 taxa) toothed whales (Odontoceti) are not related to baleen whales (Mysticeti). So the traditional clade ‘Cetacea’ is invalid or a junior synonym for Placentalia (sans Carnviora). So let’s not compare extinct ?toothed whales (the skull is missing, but see below), like Perucetus (Fig 1, MUSM 3248), to baleen whales, like Balaenoptera.
“Here we describe Perucetus colossus—a basilosaurid whale from the middle Eocene epoch of Peru. It displays, to our knowledge, the highest degree of bone mass increase known to date, an adaptation associated with shallow diving. The estimated skeletal mass of P. colossus exceeds that of any known mammal or aquatic vertebrate”.
Are we sure this is a basilosaurid whale (Figs 1, 2)? The lumbar shapes are not elongate in Perucetus as they are in Basilosaurus. Eschrictius (Figs 1, 2) is a better match.
“We use the skeletal fraction to estimate the body mass of P. colossus, which proves to be a contender for the title of heaviest animal on record.”
The authors’ diagram (Figs 1) indicate a contender status. The bones alone (Fig 2) indicate a much smaller lumbar region, in the range of the extant gray whale.
Figure 2. Eschrichtius has a robust lumbar series, like Perucetus. Note the sacrals (orange) on Perucetus are starting to taper, which makes estimating the caudal length possible. This is much shorter than Bianucci et al guessed using a basilosaurid Bauplan.
Perucetus is from the Eocene. Eschrichtius is the extant gray whale. Both share 11 robust lumbars. The shallow ribs of Perucetus hint at a more gracile torso than the gray whale (Fig 3).
Maybe the wrong Bauplan was chosen for Perucetus. It seems to be a mid-sized mysticete with short ribs and a short tail – not a giant, fat basilosaurid. Dense bones make sense for a gray whale-like mysticete that may also have spent time stirring up sea sand to filter feed.
Alas, the skull remains unknown. Even so, since mysticetes and odontocetes evolved separately, we don’t always have to look to basilosaurids for a primitive Bauplan. Build your own LRT to find this out for yourself.
PS Could Perucetus have been transitional between desmostylians and mysticetes? According to Hayashi et al 2013, “Desmostylians are, with cetaceans, the second mammal group showing a shift from bone mass increase to a spongy inner organization of bones in their evolutionary history.”
So, vertebral bone density in Perucetus is a unique, derived trait. Hippos have dense limb bones, which enables them to walk and gallop underwater on the bottom of the lake
Taxa that should be considered include two pre-desmostylians, Janjucetus and Mammalodon, both known chiefly from skull material both estimated at 3m in length.
References Bianucci G et al (15 co-authors) 2023. A heavyweight early whale pushes the boundaries of vertebrate morphology. Nature. doi:10.1038/s41586-023-06381-1 Hayashi S et al (8 co-authors) 2013. Bone Inner Structure Suggests Increasing Aquatic Adaptations in Desmostylia (Mammalia, Afrotheria). PlosOne https://doi.org/10.1371/journal.pone.0059146
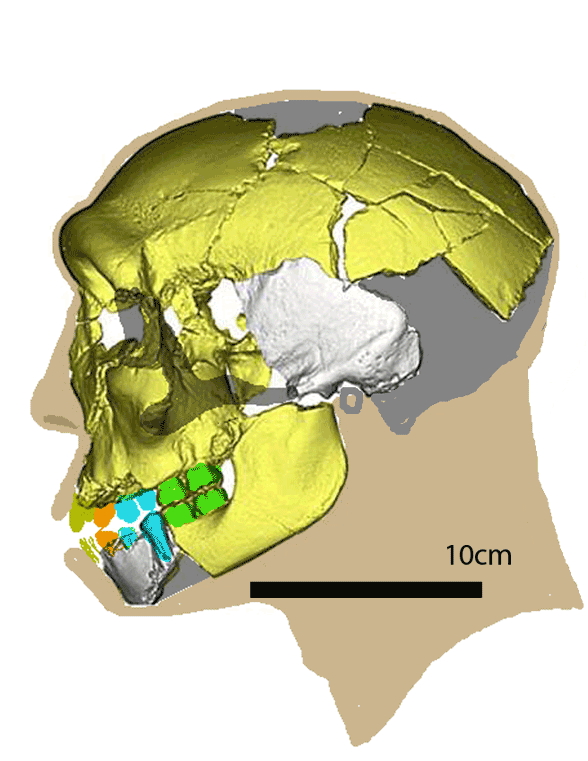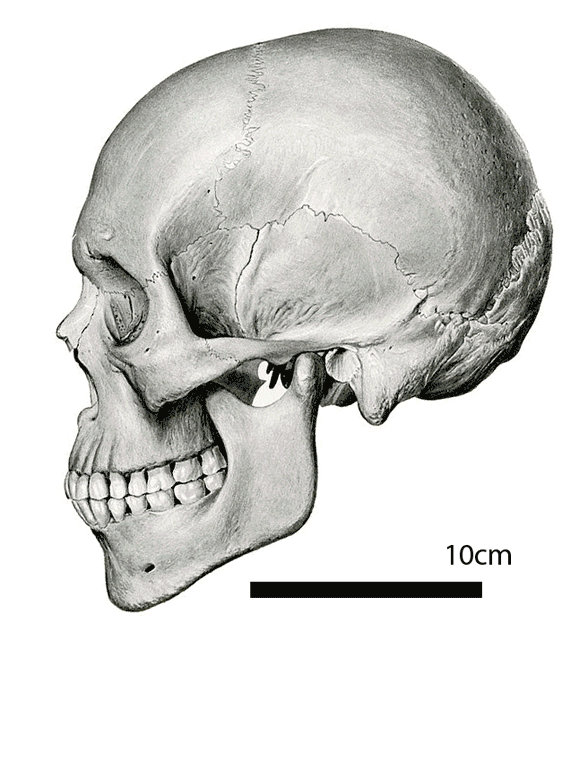
Wu et al 2019, 2023 are piercing together Middle Pleistocene (300,000ya) hominid remains (Fig 1, 2) that remained unnamed. The museum number is HLD6.
Figure 1. HLD6 skull in two views. Teeth and missing parts restored here.
Wu et al 2923 report, “Thus far, detailed morphological description and comparisons of the mandible have not been conducted. Here we present a comprehensive morphological, metric, and geometric morphometric assessment of this mandible and compare it with both adult and immature specimens of Pleistocene hominins and recent modern humans. The weak expression of all these features indicates that this mandible does not possess a true chin. The combination of both archaic and modern human features identified in the HLD 6 mandible is unexpected. With these findings, it is possible that modern human morphologies are present as early as 300 ka and earlier than the emergence of modern humans in East Asia.”
Figure 2. HLD6 skull to scale with extant Homo sapiens skull.
From DiscoverMagazine.com “The evolution of the chin has long been debated in paleoanthropology circles, but everyone agrees only Homo sapiens have prominent ones.
“Multiple specimens of Homo erectus and related lineages have been unearthed going back more than 1.6 million years.
“The Recent African Origin (RAO) model of human evolution has dominated. According to multiregionalists, when H. erectus left Africa almost 2 million years ago, these early explorers didn’t die out. They fanned out across Eurasia and continued to evolve, eventually becoming regional populations of H. sapiens before other populations of H. sapiens left Africa.
“In the last two years, some researchers have proposed a third model that tries to reconcile the dueling models and the expanding fossil record, acknowledging the complexity of our origin story. At about 300,000 years old, HLD 6 may be evidence of regional continuity, a transitional human that represents the evolution of archaic East Asian H. erectus populations into East Asian AMHs.”
AMH = Anatomically Modern Humans
Figure 3. The gibbon lineage leading to humans. At right is Australopithecus, a bipedal ape by convergence with humans.
Phylogenetic analysis in the Large Reptile Tree (LRT, 2295 taxa) indicates an East Asian origin for the genus Homo (Fig 1) from the gibbon (Hylobates. Fig 3) lineage. In the same analysis, Australopithecus, (Fig 3) the traditional human ancestor, nests instead with apes as a biped by convergence.
References Wu X et al 2023. Morphological and morphometric analyses of a late Middle Pleistocene hominin mandible from Hualongdong, China, Journal of Human Evolution (2023). DOI: 10.1016/j.jhevol.2023.103411 Wu X et al 2019. Archaic human remains from Hualongdong, China, and Middle Pleistocene human continuity and variation, Proceedings of the National Academy of Sciences (2019). DOI: 10.1073/pnas.1902396116
Brazilian host, Alex Kellner, begins his introductory lecture at 33:20 following instrumental music. The entire video extends to 1:30 hours.
Kellner acknowledges “tremendous amount of dissent” among pterosaur researchers. 240 species. The large pterosaur tree (LPT, 269 taxa) tests 255 pterosaurs and 14 outgroup taxa. Kellner indicates widespread rejection of bipedalism in pterosaurs, but then shows Bennett’s 1997 reconstruction of an upright Nyctosaurus (Fig 1) based on UNSM 93000.
Figure 1. Nyctosaurus reconstruction according to Bennett (1997) and Peters, both based on UNSM 93000.
Kellner wonders about pterosaur origins squamates (no one supports this, instead Kellner is referring to lepidosaurs like Huehuecuetzpalli (Fig 2) according to Peters 2007, still the only worker who has explored this possibility), basal archosaurs (lagerpetids? proterosuchids? euparkeriids? Bennett 1997), and archosaurs closer to dinosaurs (everyone else supports this, Padian 1987). Kellner cites three workers: David Hone, Mike Benton and Chris Bennett. Not sure how they match to the three theories. Actually, they don’t match at all.
Kellner suggests the reason for this lack of consensus comes down to a lack of intermediate stages in the genesis of pterosaurs. That ignores Peters 2000, 2002, 2007 that provided half a dozen intermediates (Fig 2) going back to Macrocnemus and Huehueceutzpalli (Fig 2).
Remember Bennett’s curse? “You will not be published and if you are published, you will not be cited.” Kellner’s remarks represent just the latest version of this attitude.
Figure 2. The evolution of the pterosaur tail beginning with a basal lizard, Huehuecuetzpalli.
Kellner’s pterosaur origin wish list: 1. “A piece of a pterosaur ?number [closed caption translation can’t deal with Kellner’s accent] because the whole skeleton would be, you know, too much to ask for”. 2.“I would love to find a creature that would if it could speak in English, Portuguese, whatever, would say I want to become a pterosaur when I evolve”.
That cuts straight to the issue. Professor Kellner would love to find this creature. He is not happy that an amateur discovered six such creatures in 2000 from previously published taxa. And since an amateur published on this (Kellner was a referee) he and other PhDs have not and will not examine this possibility, perferring instead to repeat textbook traditions and myths.
Kellner explains all known pterosaurs already have “the same Bauplan as a pterosaur, with a a very large fourth wing finger. a large metacarpal, a deltopectoral crest of the humerus and also the sternum.”
This is a wish list. A myth list. Kellner ignores actual evolutionary events in which the wings evolved last (Fig 2, Peters 2002), as in birds and bats. And, once again, this is why I am so disappointed with pterosaur workers and their workshops. Let’s remember that a pterosaur has a sternal complex = combined sternum, clavicles and interclavicles (Fig 3) that evolved from taxa like Huehuecuetzpalli (Fig. 2). Ask pterosaur professors why they keep their blinders on. We called this a curious lack of curiosity, especially for a scientist. It appears to come down to self-preservation and maintaining professional status, so this is human and forgivable, but at the same time, this is silly.
Figure 3. Tritosaur pectoral girdles demonstrating the evolution and migration of the sternal elements to produce a sternal complex.
Kellner than semi-embraces the novel and invalid lagerpetid hypothesis of pterosaur origins based on multi-cusped teeth (as inLongisquama, Fig 4)), brain case arguments and indeterminate lower jaw arguments. This is silly. He should be adding taxa to determine a last common ancestor in phylogenetic analysis. He should not be cherry-picking traits. This always leads to trouble.
It is notable that taxa cited in Peters 2000 are once again omitted, just as they were omitted from Hone and Benton 2009. This is not good science, but it has been acceptable science in academic circles for over two decades.
Fig. 4. A short history of Longisquama skull tracings. All workers have traced an antorbital fenestra, but Senter (2003) reported it was not present. Note the level of detail provided by the DGS method in the lower tracing.
Kellner puts the lagerpetid origin hypothesis blame on cladistics, “That’s the result and that’s what we have to accept,” he says. Curious that lagerpetid supporters are all from South America where lagerpetids are found. Despite his own doubts Kellner concludes the lagerpetid hypothesis is a breakthrough. This is bad science. Then Kellner acknowledges and dismisses Scleromochlus from Scotland (Fig 5), which he reports, “there’s not even a bone there, but there were impressions.”
Kellner otherwise ignores the issue of two decades of taxon exclusion (Fig 2).
Question for Kellner: Why even bring up the subject of pterosaur origins if you have nothing valid to base your talk on? This is called a waste of time delving into one fantasy = invalid hypothesis after another.
Question from Kellner: “How do you turn a lagerpetid into a viable pterosaur?”
Answer: you don’t. Seek your pterosaur ancestors elsewhere (Fig 2).
Answer from Kellner: “I want to see something in the hand, (Fig 5) I want to see something in their arms, the pelvis, they have a pubischtic plate.”
See figure 6.
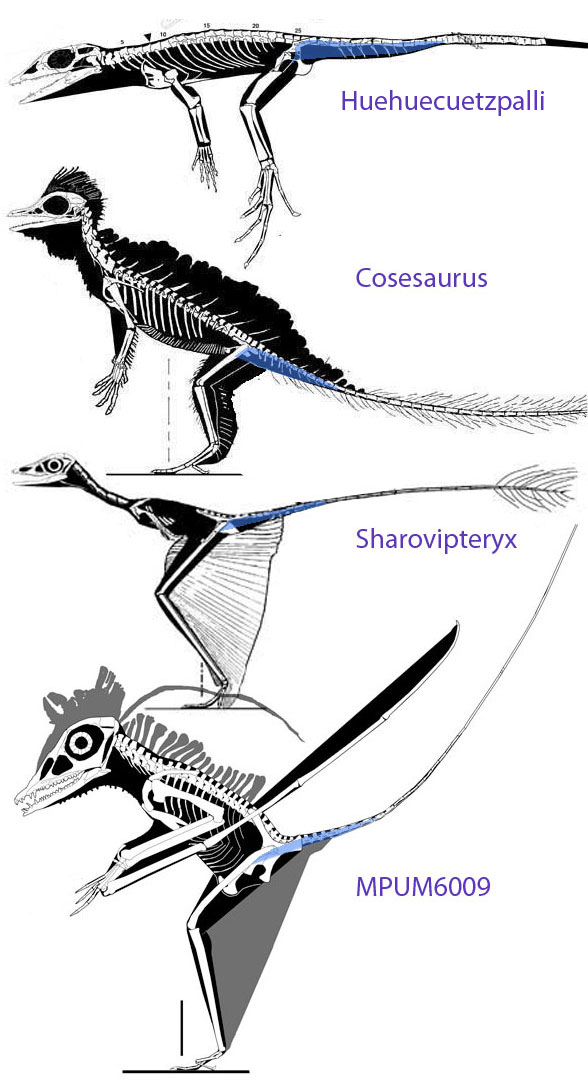
Figure 5. Click to enlarge. The origin of the pterosaur wing and the migration of the pteroid and preaxial carpal. A. Sphenodon. B. Huehuecuetzpalli. C. Cosesaurus. D. Sharovipteryx. E. Longisquama. F-H. The Milan specimen MPUM 6009, a basal pterosaur.
If Kellner is looking instead for the pterosaurian prepubis Kellner can find that in Cosesaurus, Sharovipteryx and Longisquama (see Fig 6). But he and others don’t want to go down that path.
Figure 6. Origin and evolution of the prepubis in tritosaurs.
At 50:35 Kellner blames the current phylogenetic arguments on incomplete material. This is so short-sighted. We have so many wonderfully complete specimens that never enter phylogenetic analyses, other than the LPT.
Once again, taxon exclusion, not incomplete material, is the number one issue facing paleontology in general, and specifically in pterosaurs. Workers need to build their own wide gamut phylogenetic analysis to find out for themselves how pterosaurs are interrelated. 200+ taxa is the number to shoot for.
I am so glad that I did not have to suffer under the tutelage of current professors in order to study pterosaurs and their precursors. I was lucky that Sharovipteryx and Longisquama (Figs 2–4) came to my hometown at the moment I was curious enough and ready to study them. I was lucky that Paul Ellenberger provided me with unpublished material on his little Cosesaurus. His observations enhanced my freshman studies of this important fossil wrapped in toilet paper in Barcelona. I was lucky that, like Kellner, everyone else studying pterosaurs was looking for something in the wing-finger and hand, not realizing that wings came last in pterosaur evolution (Peters 2002), as they did in birds and bats. I studied the feet first (Peters 2000a) and the rest followed. I was lucky that David Hone rejected my update on Cosesaurus and kin. That frustration with academic corruption and their lack of curiosity lead to the creation of ReptileEvolution.com and PterosaurHeresies.Wordpress.com.
Over the last 12 years that drive to learn led to the addition of 2292 taxa in the LRT plus 269 taxa in the LPT without the intervention of professional gatekeepers who continue to omit pertinent taxa in their hope to discover the origin of pterosaurs themselves. Some things cannot be discovered twice.
You can recover the origin of pterosaurs yourself by repeating this online experiment. Add taxa. That’s all takes. Let us all know if your results with a similar wide gamut taxon list confirm, refute or modify published analyses, including the traditionally ignored LPT.
References Bennett SC 1996. The phylogenetic position of the Pterosauria within the Archosauromorpha. Zoolological Journal of the Linnean Society 118: 261–308. Bennett SC 2008. Morphological evolution of the forelimb of pterosaurs: myology and function. Pp. 127–141 in E Buffetaut and DWE Hone (eds.), Flugsaurier: pterosaur papers in honour of Peter Wellnhofer. Zitteliana, B28. Carroll and Thompson 1982. A bipedal lizardlike reptile fro the Karroo. Journal of Palaeontology 56:1-10. Dalla Vecchia FM 2009. Anatomy and systematics of the pterosaur Carniadactylus (gen. n.) rosenfeldi (Dalla Vecchia, 1995). Rivista Italiana de Paleontologia e Stratigrafia 115 (2): 159-188. Ellenberger P and de Villalta JF 1974. Sur la presence d’un ancêtre probable des oiseaux dans le Muschelkalk supérieure de Catalogne (Espagne). Note preliminaire. Acta Geologica Hispanica 9, 162-168. Hone DWE and Benton MJ 2007. An evaluation of the phylogenetic relationships of the pterosaurs to the archosauromorph reptiles. Journal of Systematic Palaeontology 5:465-469. PDF online Hone DWE and Benton MJ 2008. Contrasting supertree and total-evidence methods: the origin of the pterosaurs. In: Hone DWE, Buffetaut E, editors. Flugsaurier: pterosaur papers in honour of Peter Wellnhofer. Vol. 28. Munich: Zittel B. p. 35-60. Nesbitt SJ 2011. The early evolution of archosaurs: relationships and the origin of major clades. Bulletin of the American Museum of Natural History 352: 292 pp. Nopcsa F 1931.Macrocnemus nicht Macrochemus. Centralblatt fur Mineralogie. Geologic und Palaeontologie; Stuttgart. 1931 Abt B 655–656. Peabody FE 1948. Reptile and amphibian trackways from the Lower Triassic Moenkopi formation of Arizona and Utah. University of California Publications, Bulletin of the Department of Geological Sciences 27: 295-468. Peters D 2000a. Description and Interpretation of Interphalangeal Lines in Tetrapods. Ichnos 7:11-41. Peters D 2000b. A reexamination of four prolacertiforms with implications for pterosaur phylogenesis. Rivista Italiana di Paleontologia e Stratigrafia 106: 293–336. Peters D 2002. A New Model for the Evolution of the Pterosaur Wing – with a twist. – Historical Biology 15: 277–301. Peters D 2007. The origin and radiation of the Pterosauria. In D. Hone ed. Flugsaurier. The Wellnhofer pterosaur meeting, 2007, Munich, Germany. p. 27. Peters D 2009. A reinterpretation of pteroid articulation in pterosaurs. Journal of Vertebrate Paleontology 29: 1327-1330 Renesto S and Avanzini M 2002. Skin remains in a juvenile Macrocnemus bassanii Nopsca (Reptilia, Prolacertiformes) from the Middle Triassic of Northern Italy. Jahrbuch Geologie und Paläontologie, Abhandlung 224(1):31-48. Sharov AG 1971. New flying reptiles from the Mesozoic of Kazakhstan and Kirghizia. – Transactions of the Paleontological Institute, Akademia Nauk, USSR, Moscow, 130: 104–113. Snyder RC 1954. The anatomy and function of the pelvic girdle and hind limb in lizard locomotion. American Journal of Anatomy 95:1-46 Wild R 1978. Die Flugsaurier (Reptilia, Pterosauria) aus der Oberen Trias von Cene bei Bergamo, Italien. Bolletino della Societa Paleontologica Italiana 17(2): 176–256. Wild R 1993. A juvenile specimen of Eudimorphodon ranzii Zambelli (Reptilia, Pterosauria) from the upper Triassic (Norian) of Bergamo. Rivisita Museo Civico di Scienze Naturali “E. Caffi” Bergamo 16: 95-120. pterosaurheresies.wordpress.com/2011/07/16/what-exactly-is-a-pterosaur-part-3-of-3/ reptileevolution.com/pterosaur-wings.htm
Peters D unpublished update on Cosesaurus, Sharovipteryx and Longisquama.
The base of the Porolepiformes have been problematic in the large reptile tree (LRT, 2295 taxa). Data for Quebecius comes from two vintage diagrams (Fig 1). One has a high forehead. The other has a more bullet-shaped rostrum. The fossil photo (Fig 1, upper right) seems closer to the bullet-shaped rostrum diagram (Fig 1 center).
Figure 1. Quebecius has pectoral lobe fins and pelvic ray fins. Note the two slightly different faces. Compare to figure 2.
Note the distinctive fin shapes in Quebecius (Fig 1). The pectoral fin is like a long, cone-shaped lobe, but the pelvic fin is only made of rays, almost rectangular, not lobe-shaped at all. This trait disparity sets Quebecius apart from other porolepiformes with four lobe-fins. Among other fish taxa only Cheirolepis (Fig 2) has a similar disparate fin pattern morphology, mixing pectoral lobe-fins with pelvic ray-fins.
Figure 2. Bulet-head Quebecius skull with DGS colors added and reconstructed. These skull changes link Quebecius more closely to Cheirodus, the taxon below the scale bar. Compare to figure 1.
After DGS coloring of the Quebecius skull (Fig 2) and rescoring this taxon in the LRT, Quebecius now nests with Cheirolepis. This resolves only one last lingering problem in the LRT.
Testing all fish sans sharks, placoderms and more basal chordates (329 non-deleted taxa) took only 55 seconds to fully resolve. Testing only sharks, placoderms and more basal chordates (155 taxa) takes only 02 seconds to fully resolve. Seemingly following an exponential increase, testing all fish taxa (total 484) takes much longer: 43 minutes to fully resolve.
Figure 3. Miguashaia juvenile and adult. These are lobe-fin porolepiformes. Note the pectoral fins.
Quebecius quebecensis (Hay 1902, originally Holoptychius quebecensis Whiteaves 1889; Schultze and Arenault 1987; Late Devonian, up to 60cm in length, Figs 1, 2) was originally considered a porolepiform, but here the bullet-skull specimen nests with Cheiroleips (Fig 2). Like Cheirolepis, the fore fin is lobed, but the pelvic fin is not. These traits pull Quebecius away from Miguashaia(Fig 3), Holoptychius and other lobe-finned taxa in the Porolepiformes.
References Hay OP 1902. Bibliography and Catalogue of the Fossil Vertebrata of North America. Bulletin of the United States Geological Survey 179:1-868. Whiteaves JF 1889. -pt. III. Fossils of the Devonian rocks of the Mackenzie River basin. Contributions to Canadian Paleontology, Geological Survey of Canada.