
Gavazzi et al. 2020 bring us their views
on whale and pre-whale carpal elements. They used the invalidated artiodactyls, pig (Sus) and (Diacodexis) as outgroup taxa. The latter is incompletely known, slender and appears to be not far from the more completely known Cainotherium.
By contrast
here (Figs. 1–3, 5, 7, 8) competing whale and pre-whale carpal elements are presented along with tree shrew (Ptilocercus) carpals (Fig. 4) as an example of the plesiomorphic condition in placentals.
Bottom line:
Due to taxon exclusion, actual evolutionary patterns were overlooked by Gavazzi et al., but that didn’t matter much. About the same story can be told with the wrong outgroup taxa. Carpals don’t change much across these members of the placental clade.

Figure 1. Odontocete flipper and ancestral taxa manus. Homologous wrist elements are colored the same. Green is the pisiform, missing in the dolphin, Tursips. Ambulocetus image from Gavazzi et al. 2020 and repaired here. Dorudon image from Cooper et al. 2007 and repaired here.
From the Gavazzi et al. 2020 abstract:
“During the land-to-water transition in the Eocene epoch, the cetacean skeleton underwent modifications to accommodate life in the seas. These changes are well-documented in the fossil record. The forelimb transformed from a weight-bearing limb with mobile joints to a flipper with an immobile carpus.”
Unfortunately Gavazzi et al. still consider ‘Cetacea’ a monophyletic clade. That hypothesis of interrelationships became invalid in 2017 with a blogpost here and an online paper here.

Figure 2. The manus of Eubaelana, an extant right whale (bottom) compared to two desmostylians. Note the similarities between the two unrelated clades. Missing parts are restored based on phylogenetic bracketing.
Mammal carpal homologs:
- Distal carpal 1 = Trapezium – red
- Distal carpal 2 = Trapezoid – cyan
- Distal carpal 3 = Capitum – lavender
- Distal carpal 4+5 = Hamatum – yellow green
- Radiale = Scaphoideum – pink
- Intermedium = Lunatum – yellow
- Ulnare = Triquetrum – pale orange
- Centrale = Centrale – indigo
- Pisiform = Pisiform – green

Figure 3. Hippos manus. Pisiform in green.
Continuing from the Gavazzi et al. 2020 abstract:
“We used micro-CT imaging to assess evolutionary changes in carpal size, orientation, and articulation within Eocene cetacean taxa associated with the transition from a terrestrial to amphibious niche. We compared Ambulocetus natans (Fig. 1), a well-preserved amphibious archaeocete, with other archaeocetes, and with Eocene terrestrial artiodactyls, the sister group to Cetacea.”

Figure 4. Ptilocercus is a tree shrew closer to the plesiomorphic wrist condition. Compare to taxa in figures 1 and 2.

Figure 5. X-ray of Tursiops flipper showing no trace of a pisiform. But look at digit 5. Compare to figure 4.
Eocene terrestrial artiodactyls are not the sister group to Cetacea, which is not a monophyletic clade in the LRT. Adding overlooked taxa resolves this issue.
“A cylindrical carpus in terrestrial taxa evolved into a mediolaterally flattened, cambered carpus in the semi-aquatic and fully aquatic cetaceans.
Gavazzi et al. chose the wrong terrestrial taxa. Tenrec carpals (Figs. 1, 7) are not as cylindrical as those of pigs and their ancestors, though the pisiform does orient itself ventroposteriorly. All flippers are flattened with a lateral pisiform, which turns out to be a reversal back to the tree shrew orientation (Fig. 4).
“Specifically, the pisiform bone shifted from a ventral [= posterior, Fig. 3] orientation in terrestrial taxa to a lateral orientation, in plane with the carpus, within semi-aquatic and fully aquatic taxa.”
A ventroposterior pisiform is also found in Tenrec (Fig. 1) and Hippopotamus (Fig. 3) among the actual ancestors of odontocetes and mysticetes respectively.
“Flattening of the carpus, including lateral rotation of the pisiform, likely relates to functional shifts from weight-bearing terrestrial locomotion to aquatic locomotion. This laterally projecting pisiform morphology is retained in all extant cetaceans.”
Gavazzi et al. used the wrong outgroup taxa. In both Tenrec and Hippopotamus the carpus is less cylindrical than in the artiodactyls, Sus and Diacodexis.
By the way, the pisiform is absent in the dolphin Tursiops (Figs. 1, 5), but look at the lateral orientation of digit 5 in comparison to the lateral orientation of the pisiform in the plesiomorphic wrist of Ptilocercus (Fig. 4). One wonders if Gavazzi et al. mistook digit 5 in Tursiops for a pisiform, given their statement.
“Our results suggest this shift, along with other modifications to the carpus, predominantly occurred during the middle Eocene and facilitated an obligatorily aquatic lifestyle in late Eocene cetaceans.”
Without the real ancestors, the Gavazzi et al. paper and its conclusions would be a waste of time, except that carpals are largely interchangeable in quadrupedal placentals. So their conclusions remain largely valid. Thirteen years ago Cooper et al. 2007 also looked at whale carpals and compared them to predecessor taxa (see below).
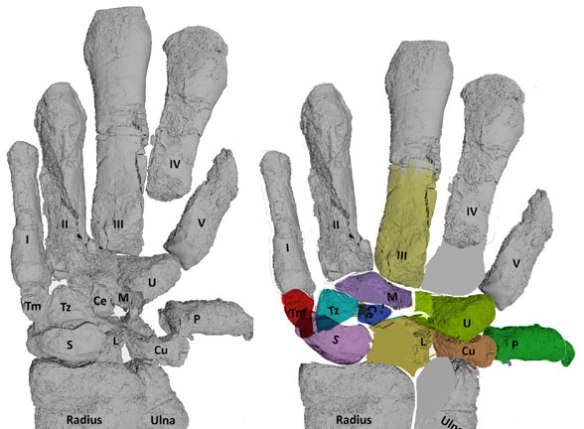
Figure 6. Ambulocetus carpals from Gavazzi et al. (left) here restored to their original positions and colored.
Try to always restore scattered elements
to their original positions. Don’t leave the work half done. Colors help to make this process easier and aid in making confident comparisons between taxa.

Figure 7. Manus and wrist of the extant tenrec Hemicentetes (from Digimorph.org and used with permission). Colors added. Compare to related taxa in figures 1 and 6. Pisiform in bright green.
The traditionally overlooked overall resemblance of tenrecs
to Indohyus (Fig. 8) and later odontocete ancestors (Fig. 8) extends to the manus and carpus.
Whale workers who refereed the manuscript
‘The Triple Origin of Whales‘ declined to allow it to be published. In that light, one wonders why whale workers prefer to cherry-pick pigs and pig ancestors rather than adding valid taxa to their cladograms. Keeping their blinders on is a continuing problem in paleontology, which is why this blogpost exists. For those readers hoping someday to make discoveries in this field of science, beware. Many professors will attempt to suppress your work to keep the discoveries for themselves.

Figure 8. Odontoceti (toothed whale) origin and evolution. Here Anagale, Andrewsarchus, Sinonyx, Hemicentetes, Tenrec Indohyus and Leptictidium precede Pakicetus. Maiacetus and Orcinus are aquatic odontocetes.

Figure 9. Taxa in the lineage of right whales include Desmostylus, Caperea and Eubalaena. The tiny bit of jugal posterior to the orbit (in cyan) is found in all baleen whales tested so far. The frontals over the eyes are just roofing the eyeballs in Desmostylus, much wider in Caperea and much, much longer in Eubalaena.
Cooper et al. 2007
dissected extant whale flippers and compared them to Ambulocetus (Fig. 1) and Dorudon (Fig. 1) without correcting for disarticulation problems in fossils. Cooper et al. report, “Most odontocetes also reduce the number of phalangeal elements in digit V, while mysticetes typically retain the plesiomorphic condition of three phalanges.” Perhaps Cooper et al. did not notice there are four phalanges on digit five in Eubaelana (Fig. 2). The unguals are tiny.
References
Cooper LN, Berta A, Dawson SD and Reidenberg JS 2007. Evolution of hyperphalangy and digit reduction in the cetacean manus. The Anatomical Record Special Issue: Anatomical Adaptations of Aquatic Mammals 290(6): https://doi.org/10.1002/ar.20532
Gavazzi LM, Cooper LN, Fish FE, Hussain ST and Thewissen JGM 2020. Carpal Morphology and Function in the Earliest Cetaceans. Journal of Vertebrate Paleontology. DOI: 10.1080/02724634.2020.1833019
https://www.researchgate.net/publication/328388746_The_triple_origin_of_whales

Dear mr/dr Peters,
I have been reading your blog for a few months, and I have a deep interest in the pterosaur question. Not being an expert in any way, I have really learnt from your simple, forthright explanations to some interesting fossils and concepts. Recently I read some rather awful statements from various other people in your field, and I would just like to say that, you have been maligned for difference of opinion. This is wrong. The field cannot move forward without some “ blue sky thinking”. I’m sorry that you quest to illuminate is being smothered by the very system that supports it, but I and a few friends relish the insights. My friends and I are English, and from the north, so we expect and deal with monopoly and “Pooh-poohing”. Keep it up , and a you admiral farragut said “full speed ahead, damn the torpedoes!”
Stuart cook
Thank you, Stuart. As you note, I never thought science would be like this, especially when the antagonists are teachers.