Short one today
as the graphics (Fig 1) tell the story recovered by the large reptile tree (LRT, 2299 taxa).


Short one today
as the graphics (Fig 1) tell the story recovered by the large reptile tree (LRT, 2299 taxa).
A 1m long Amazon fish nicknamed ‘water monkey’
for its ability to pick vertebrate prey off low-hanging branches, Osteoglossum (Fig 1), now has a North American relative, the 45cm goldeye (Hiodon alosoides, Fig 2) in the large reptile tree (LRT, 2299 taxa).
These two are both freshwater taxa and are not traditionally linked together.

Along similar lines
the 13cm West African butterflyfish, Pantodon (Fig 2) also has a North American relative in the LRT, the 45cm mooneye, Hiodon tergisus (Fig 2).
These two are both freshwater taxa and are not traditionally linked together.
Common ancestors of all of these taxa
must have existed prior to the appearance of the Atlantic Ocean during the splitting of North and South America from Africa. Either that or these freshwater taxa traversed the salty Atlantic.

Osteoglossum and Pantodon
are traditional members of the Osteoglossiformes, named for their bony tongue. Hiodon alosoides also has a bony tongue, labeled by Gregory 1933 (Fig 3) as a glossohyal plate.

According to Wikipedia
“the freshwater butterflyfish or African butterflyfish, Pantodon buchholzi, is the only extant species in the family Pantodontidae within the order Osteoglossiformes.”
In the LRT every tested taxa is related to every other tested taxa.
The only question is “what is the proximity?”
According the the LRT
Pantodon is more closely related to Hiodon tergisus while Osteoglossum is more closely related to Hiodon alosoides. These hypothetical interrelationships call into question the genus Hiodon and the clade Osteoglossiformes.
Minor discoveries like these are the rewards that come with building your own LRT.
If you haven’t started one yet, it’s time to. Or you can just rely on out-dated textbooks.

This appears to be a novel hypothesis of interrelationships.
If not, please provide a citation so I can promote it here.
References
wiki/Goldeye
wiki/Osteoglossum
wiki/Mooneye
wiki/Freshwater_butterflyfish
This sometimes giant, usually lethargic Amazon River fish
has an armored heavily textured skull (Fig 1, Arapaima gigas) that is newly interpreted with DGS colors to match the Bauplan of the extant walking catfish, Clarias (Fig 2) one of the most primitive bony ray-fin fish in the LRT.


This new interpretation
solves several long-standing phylogenetic problems. And it opens the door to new hypotheses of interrelationship adding one more origin story to the operculum and preoperculum in gnathostomes.

Small steps like this one
(Fig 1) are slowly rebuilding the fish subset of the large reptile tree (LRT, 2299 taxa). It’s a learning experience.
Bonus YouTube Video featuring PaleoArtist Mark Hallett
Sometimes it helps to add taxa.
The software can only work with what you give it. And you have to be willing to test older results that appear to have merit… but don’t quite match.
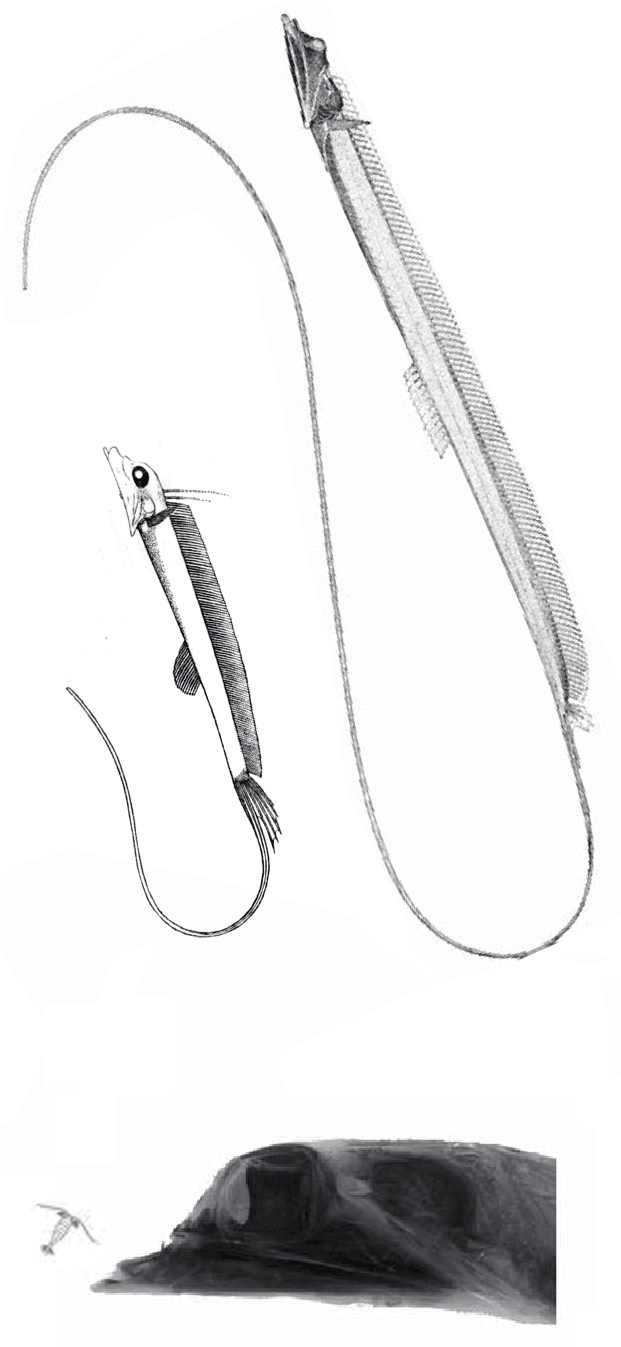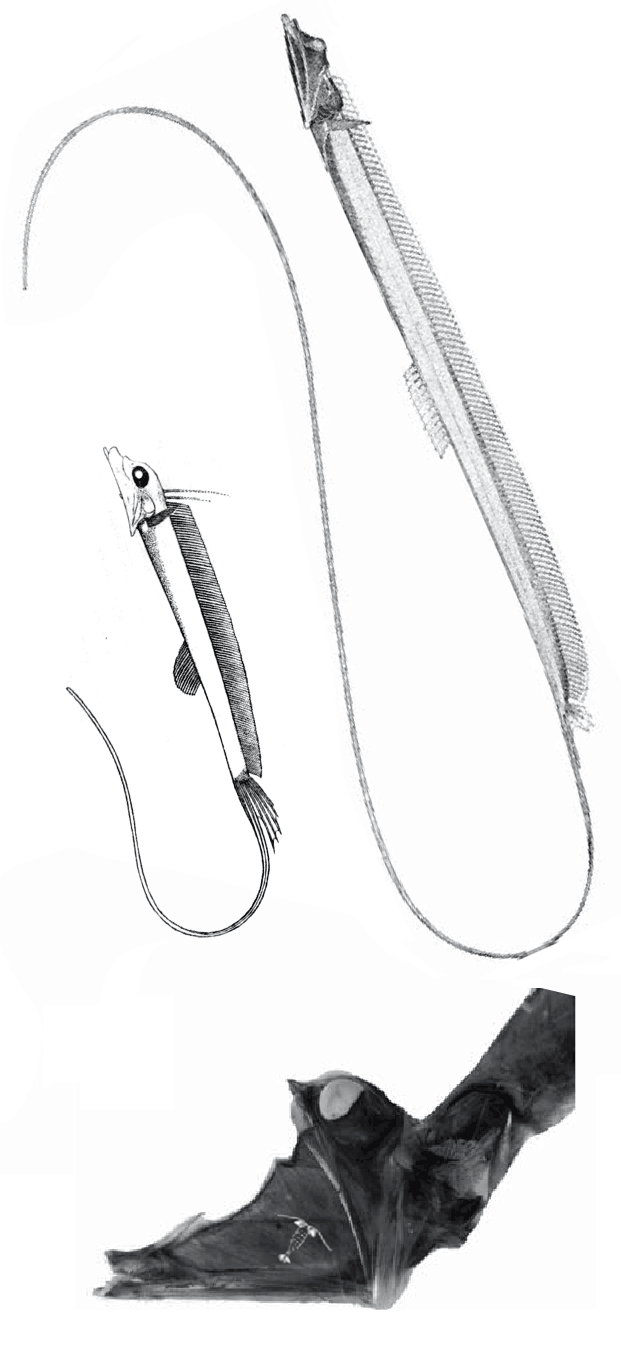
Stylephorus chordatus
(Shaw, 1791, Regan 1924) is the extant tube-eye or thread-tail. It was considered an oarfish relative, but here nests with Spinocaudicthys (Fig 2). Note the large eyes and flexible neck. Like seahorses and oarfish, the tube-eye feeds on tiny plankton sucked in as its tubular mouth enlarges the oral cavity by 40x.

Spinocaudichthys oumtkoutensi
(Filleul A and Dutheil DB 2001, Poi-SGM 20a, 3.5cm long, Cenomanian, Early Cretaceous) was originally considered a tiny acanthomorph, but in the LRT nests with extant sticklebacks, like Gasterosteus.

According to Wikipedia (Acanthomorpha)
Stylephorus (= Stylephoriformes) is most closely related to codfish (Gadus). The LRT does not support that hypothetical interrelationship.
Accordlng to Wikipedia (Sticklebacks)
are most closely related to scorpionfish, like Pterois. In the LRT sticklebacks and their kin split off from the base of the scorpionfish clade.

References
Filleul A and Dutheil DB 2001. Spinocaudichthys oumtkoutensis a freshwater acanthomorph from the Cenomanian Morocco. Journal of Vertebrate Paleontology 21(4):774–780.
Shaw G 1791. Description of the Stylephorus chordatus, a new fish. Transactions of the Linnean Society of London, 2d Ser: Zoology 1:90–92.
wiki/Stylephorus
wiki/Spinocaudichthys – not yet posted
For years, I found it difficult to understand
the naris and nares of the extant anchovy, Engraulis (Fig 1). A small hole at the snout tip appears to be an incurrent naris. Then, working slightly backwards, things get a little confusing looking for the excurrent naris hole or slit.

Comparisons to Late Triassic Birgeria
(Fig 2) were as instructive as confusing.

Recent comparisons to Late Devonian Gogosardina
(Fig 3) finally helped understand homologies… IF facial bones were permitted to split and fuse, as they often do in fish. See the split happen in the two frames of this GIF animation.

Engraulis encrasicholus (Linneaus 1758; up to 21cm) is the extant European anchovy. As a primitive teleost, the jaw joint is behind the orbit. Fast swimming anchovies travel in open water schools.
Birgeria stensioei (Agassiz 1844; Romano and Brinkman 2009; Late Triassic; 1-2m, PIMUZ T4780) is a large, carnivorous sister to Kalops. The palatine teeth were larger than the marginal maxillary teeth.
Gogosardina coatesi (Choo, Long and Trinajstic 2009; Late Devonian) The skull is nearly identical to coeval and similarly-size Mimipiscis with slightly rotated premaxilla, a straighter anterior maxilla, a higher naris and no complete ‘razor back’ ridge, only a partial. The skull is proportionally larger as well. Both have a large pineal opening between the frontals. The excurrent naris is confluent with the orbit. This clade lacks postparietals. The supratemporals meet at the midline.
Occasional insights like this
help link taxa together, help identify skull bones, and help correct scores in the LRT. Whether or not this hypothesis of identification (Fig 3) is correct or not will be tested from here on out.
References
Choo B, Long JA and Trinajstic K 2009. A new genus and species of basal actinopterygian fish from the Upper Devonian Gogo formation of Western Australia. Acta Zoologica (Stockholm) 90 (Supp 1):194–210.
Linnaeus C 1758. Systema naturæ per regna tria naturæ, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Tomus I. Editio decima, reformata.
Updated October 3, 2023
with new identities for several skull bones.
This phylogenetic pairing escaped notice earlier
because I had misidentified certain skull bones and no dorsal view of the ANU V-244 specimen (Fig 1) was published. A ventral view of the flying gurnard, Dactylopterus, would have helped as well, to compare to the ventral view of the tiny placoderm.
Color-coding helps. This is what the DGS method was made for.

Placoderms are supposed to be extinct.
That hypothesis seems to be reversing itself in the large reptile tree (LRT, 2300 taxa) as housekeeping continues to reveal overlooked close relatives. The data (Fig 1) is intriguing, hopefully others will find these interrelationships worth testing.
Dactylopterus volitans (Linneaus 1758; 50 cm) is the extant flying gurnard, a bottom-feeder living in warm shallow seas.
ANU V244 (Young, Lelièvre and Goujet 2001; Hu, Lu and Young 2017; Early Devonian) is a small placoderm represented by a 3D skull that has been taken apart by the second set of authors.
References
Hu Y, Lu J and Young 2017. New findings in a 400 million-year-old Devonian placoderm shed light on jaw structure and function in basal gnathostomes. Nature Scientific Reports 7: 7813 DOI:10.1038/s41598-017-07674-y
Linneaus C von 1766. Systema naturæ per regna tria naturæ, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Tomus I. Editio duodecima, reformata. pp. 1–532. Holmiæ. (Salvius).
Young G C, Lelièvre H and Goujet D 2001. Primitive jaw structure in an articulated brachythoracid arthrodire (placoderm fish; Early Devonian) from southeastern Australia. J. Vertebr. Paleontol. 21, 670–678.
Short one today.
The graphics (Figs 1, 2, 3) tell the tale.

According to Wikipedia,
“Lepidotidae is an extinct family of fish, known from the Jurassic and Cretaceous periods. Cladistic analysis has indicated that they are close relatives of gars, with both being members of the order Lepisosteiformes. Members of the family are known from both marine and freshwater environments. Their bulky body morphology and small median fins suggests that they were slow-moving fish that were capable of performing fine movements in order to grasp prey.”

Xu 2020 wrote,
“Neopterygii is the largest known group of ray-finned fishes today, and the Luganoiidae,
recognized by its specialized skull and greatly deepened flank scales, is a stem lineage of this
group in the Middle Triassic.”
The extant armored catfish, Hoplosternum (Figs 1–3), shares ‘greatly deepened flank scales’ and many other traits, but was not mentioned by Xu 2020.

Near and Thacker 2023
recently published online a preprint on the phylogeny of the traditional clade Actinopterygii. Unfortunately the outgroup taxon for their list of extant taxa, the lungfish Polypterus, was cherry-picked by tradition and textbooks rather than selected by software from a wider gamut of primitive candidates. That means the rest of their analysis is a house of cards, ready to fall apart at the first minor tremor.
Never cherry-pick your outgroup taxon. Always let your software tell you the appropriate outgroup for your study no matter what professors teach from out-dated textbooks at the university level. Create your own LRT to find this out for yourself.
Near and Thacker also put their faith in deep time genomics. Avoid that.
References
Nearn TJ and Thacker CE in press. Phylogenetic classification of living and fossil
ray-finned fishes (Actinopterygii). Bulletin of the Peabody Museum of Natural History 65. 807ms pp. https://zenodo.org/record/8352027
Xu G-H 2020. A new species of Luganoia (Luganoiidae, Neopterygii) from the Middle Triassic Xingyi Biota, Guizhou, China. Vertebrata PalAsiatica 58(4):267–282.
wiki/Lepidotes
wiki/Semionotus
wiki/Lepidosteus
wiki/Scheenstia
wiki/Luganoia
wiki/Cladistia
wiki/Actinopteri
wiki/Lepidotidae
The 69th Symposium of Vertebrate Palaeontology and Comparative Anatomy (SVPCA) and the 28th meeting of the Symposium of Palaeontological Preparation and Conservation (SPPC) was held on 6th to 9th September 2023 at the University of Lincoln, in the Minerva Building.
You can view the SVPCA abstracts for 2023
online at their website here. Only a few merit confirmation, refutation or modification based on phylogeny and other topics generally covered here.

Unwin D et al 2023. Rhamphorhynchus: a model for ontogenetic niche partitioning in pterosaurs?
“Ontogenetic niche partitioning (ONP) has recently been proposed for several pterosaurs, primarily on the basis of size distributions that, while suggestive, are not conclusive. Resolving this question is important as ONP has potentially profound implications for the taxonomy, systematics, ecology and evolutionary history of pterosaurs. This study focused on Rhamphorhynchus muensteri, utilised a large data set (130+ individuals) that represent an almost complete growth sequence [continued below].
Unwin does not mention creating a phylogenetic analysis to lump parents with juveniles and separate small species from large species. In 2015 only one juvenile Rhamphorhynchus (Fig 3) was recovered after phylogenetic analysis of several dozen specimens (Fig 1). You can read about that hypothesis here and see reference list below for a ResearchGate PDF.
[continued] and aimed to identify putative ontogenetic niches using multiple independent data sets: comparative morphology, morphometrics and tooth microwear.
What you don’t see on Unwin’s short list: phylogenetic analysis.
[continued] Three distinct ontogenetic niches were identified: small, highly immature, flight capable invertivores (wingspan 0.3-0.4 m; <1 year old); medium-sized aerial piscivores (wingspan 0.75-1.25 m; ~2 to 7 years old); and large, osteologically mature individuals (wingspans 1.25-1.9 m; >7 years old) that seemingly represent an extension of aerial piscivory to include prey of relatively large size.
Phylogenetic analysis also recovered at least three sizes, but the order is not ontogenetic (Fig 1). That’s why it is so important to run the numbers. Unwin is assuming small taxa are young taxa. This is incorrect and so his team’s conclusions are based on an error.
Key evidence such as tooth microwear, which directly reflects changes in prey preferences, skull shape and wing-configuration are strongly correlated, permitting a high degree of confidence in the identification of ONP in Rhamphorhynchus. This pterosaur could thus be of great utility for assessing ONP in other taxa and a preliminary analysis suggests that this phenomenon is widespread in pterosaurs.”

This ONP hypothesis
had a recent popularity spurt with Tyrannosaurus, which begins life too small to conquer adult Triceratops. Unwin’s team reached their intended goal, but left the ball in the middle of the field. First determine phylogeny. Ontogeny will appear in pterosaurs as small taxa identical to large taxa following isometric lepidosaurian growth strategies. Doing what Unwin did enforces what they teach at his university: an allometric archosaurian growth strategy for pterosaurs. This is falsified by Pterodaustro and Zhejiangopterus (Fig 2) growth series, plus all three full-term pterosaur embryos.
If Unwin never mentioned ontogeny his team could have come to the same conclusion of niche partitioning based on speciation.

Evidently it’s important
for Dr Unwin NOT to run that phylogenetic analysis. That would have been a great opportunity for him to confirm, refute or modify the large pterosaur tree (LPT), which is more than a decade old now. Testing is what scientists do, but not if they think their job or reputation will suffer if their results confirm those of home-schooled scientists. It happened before.
References
Peters D unpublished. First juvenile Rhamphorhynchus reovered by phylogenetic analysis. researchgate.net/publication
Large Pterosaur Tree (LPT)
In the LRT sturgeons appeared before the advent
of the gnathostome jaws that we inherited from placoderms. Sturgeons feed by extending a ventral tube that is reinforced with bony elements (Fig 1). So it was only natural for the first naturalists to label these bones with their nearest tetrapod and gnathostome homologs.
That should stop now (Fig 1) because SOME of those elements are not homologous with gnathostome mandible elements. Better to provide certain sturgeon mandible bones with the prefix “pseudo-” in order differentiate them from gnathostome inventions. This will clarify scoring in fish cladograms.

Traditionally
Gonorhynchus was allied with Chanos, the milkfish (Fig 2) which entered the large reptile tree (LRT, 2301 taxa) today. As you’ll note, Chanos has a traditional mandible, not a ventral oral tube, like sturgeons have. Chanos nests between Albula, the milkfish, and Opsithoproctus, the deep-sea barrel-eye (Fig 2).
Today’s LRT also needs updating, but that will have to wait as problems like these arise and solutions are provided.

Chanos chanos
(Lacepede 1803, up to 1.8m in length, most 1m in length) is the extant milkfish. The head is small relative to the body and the toothless mouth is small relative to the skull. This streamlined fish is an open seas schooling speedster feeding on algae and tiny invertebrates. Each spawning produces 5 million siblings. Fossils extend back to the Cretaceous.
References
Lacepède BGE 1803. Histoire Naturelle des Poissons. Vol. 5. Chez Plassan, Imprimeur-Libraire, Paris. 803 pp.
wiki/Milkfish
wiki/Gonorynchus_gonorynchus
Yes. And No.
In this hypothesis of interrelationships the large reptile tree (LRT, 2300 taxa) lumps Malapterurus, the electric catfish, with the tiny parasitic catfish, Vandellia (Fig 3) and another long torso catfish, Ictalurus (Fig 4), the channel catfish. The electric eel, Electrophorus (Fig 2) nests nearby.

Malapterurus electricus
(Lacepede 1803, originally Silurus electricus Gmelin 1789, Murray AM and Holmes RB 2022, 1.2m in length) is the extant electric catfish, native to the fresh waters of Africa. Here it nests basal to the South American electric eel and knifefish, but closer to the tiny South American catfish, Vandellia. Like its relatives, the pectoral muscle surrounds most of the body and generates up to 400V. The skull is wider than relatives and has a sagittal crest. The premaxilla is relatively large because it is the result of fusion of the two loose parts. Note the posterior placement of the pelvic fins.



If you didn’t yet notice,
these two freshwater taxa Fig 1, 2) support the theory of continental drift with a phylogenetic split sometime before the geologic split at the genesis of the Atlantic Ocean in the Early Cretaceous, 140 mya. Catfish ancestors go back to Late Silurian placoderms.
References
Gmelin JF 1789. Caroli a Linné … Systema Naturae per Regna Tria Naturae, Secundum Classes, Ordines, Genera, Species, Cum Characteribus, Differentiis, Synonymis, Locis. Editio Decima Tertia, Aucta, Reformata/ cura Jo. Frid. Gmelin. Volume 1, part 3. Lipsiae: Impensis Georg. Emanuel. Beer.
Lacepède BGE 1803. Histoire Naturelle des Poissons. Vol. 5. Chez Plassan, Imprimeur-Libraire, Paris. 803 pp.
Murray AM and Holmes RB 2022. Osteology of the cranium and Weberian apparatus
of African catfish families (Teleostei: Ostariophysi: Siluriformes) with an assessment of genera from the Palaeogene of Africa. Vertebrate Anatomy Morphology Palaeontology 9:156–191/
