
Added September 21, 2020:
Think about a bubble net, as in humpback whales, coming form the long, dead=air storage vessel that is that elongate trachea. That long neck rotating like an inverted cone to surround confused fish just above the jaws.
Beard and Furrer 2017 conclude (or do they?)
that Tanystropheus (Figs. 1–3) was likely terrestrial.
From the abstract
“The Middle Triassic protorosaur Tanystropheus has been considered as both a terrestrial and aquatic taxon based on several lines of biomechanical and distributional evidence, but determining conclusively which habitat was most likely has remained problematic. The preservation of Tanystropheus was found to be more similar to Macrocnemus than Serpianosaurus implying carcasses of Tanystropheus originated in terrestrial or at least marginal and near-shore, shallow marine settings. That these were also the most probable habitats in life is supported by the relatively lower number of Tanystropheus (and also Macrocnemus) compared to Serpianosaurus.”

Figure 1. Tanystropheus in a vertical strike elevating the neck and raising its blood pressure in order to keep circulation around its brain and another system to keep blood from pooling in its hind limb and tail.
Unfortunately,
Tanystropheus was not a protorosaur, nor a member of the Archosauromorpha. It was a tritosaur lepidosaur as taxon inclusion would have informed the authors.
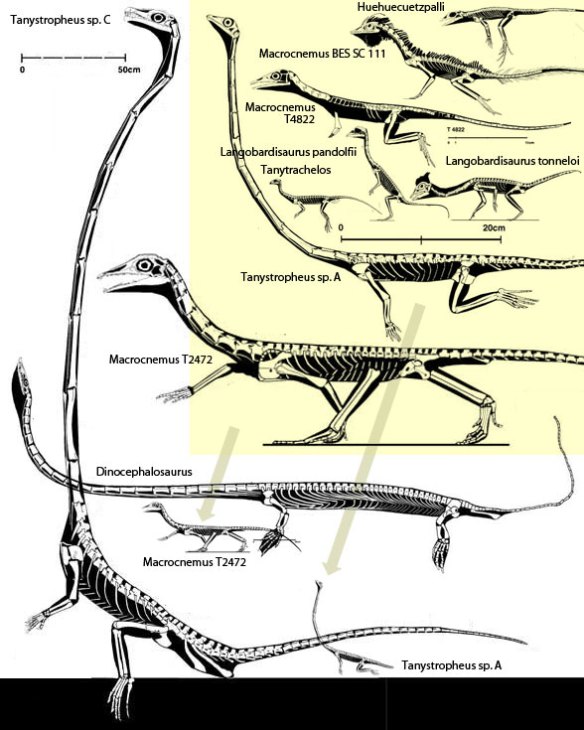
Figure 2. Tanystropheus and kin going back to Huehuecuetzpalli.
Unfortunately,
the smaller Tanystropheus considered a juvenile by the authors was probably a different genus, based on a long list of distinct traits, including its distinct teeth. Moreover the authors did not realize that several large putative Tanystropheus specimens have distinct skull morphologies that are not congeneric (Fig. 2). But all that is beside the point…

Figure 3. Tanystropheus with skull reconstructions based on two specimens, exemplar i and exemplar m.
The authors report:
“An alternative aquatic lifestyle has also been suggested for Tanystropheus (Tschanz 1988). The main argument is the supposed inflexibility of the neck due to the elongated vertebrae and bundled cervical ribs that prevented all but a horizontal position (Tschanz 1988; Renesto 2005).”
Or any aquatic position,
including vertical (Fig. 1). The authors do cite the hooks from squid suckers found in the stomach region (Wild 1973; Fig. 1) and other prior hypotheses, then describe the taphonomy of the skeletons. The authors cite trackway data matched to Tanystropheus, (Fig. 3) ignoring the fact that even sea turtles leave trackways on beaches when they lay eggs.
Figure 3. Tanystropheus specimens matched to Synaptichnium tracks. The match is good in each case, except for one toe in each trackway. So, is this good enough? Or is this cause for dismissal?
Did the authors test the tracks?
No. But that is done here (Fig. 3). In each case there is a pretty good match—except for one toe in each case. The manus and pes have been scaled to match the tracks and thus are not matched to the scale bars which are for the tracks alone. Even so, the scale for the trackmakers’ extremities is a pretty good match! The case is not rock solid, but pretty good, that big and small tanystropheids made those Synaptichnium tracks.
Taphonomy
The journey from the biosphere to the lithosphere was investigated. The terrestrial Ticinosuchus, a type of archosauriform, was discovered in these beds along with the aquatic Serpianosaurus, a type of pachypleurosaur, according to the authors (a basal thalattosaur in the large reptile tree). So was the tritosaur lepidosaur, Macrocnemus. The authors wondered if the taphonomy of Tanystropheus would be more similar to the terrestrial or the aquatic taxa. Wild (1973) listed and illustrated over a dozen specimens of Tanstropheus in various stages of completeness and articulation. The water depth of fossil deposition was estimated between 30 and 130m with anoxic bottom conditions. So ALL the specimens were transported horizontally and vertically. Importantly, fragmentary skeletons of Tanystropheus were excluded from this study. From the remaining data the authors compiled articulation and completeness scores.
Why did they throw out competing data?
We’ve seen this before with Hone and Benton (2007, 2008). Given that they used only the more complete specimens (and who knows how many incomplete specimens were never collected), the authors report Tanystropheus specimens exhibited 0-58% articulation and 36–97% completeness. Larger specimens tended to be more complete. The authors also note that Serpianosaurus alone lacks obvious features that promoted buoyancy, like hollow cervicals in Tanystropheus. The authors cite Brand et al. 2003, who noted “lizard skin in water formed a limp but durable bag containing the bones” during water transport.
The authors conclude
“Tanystropheus langobardicus at least died in, but probably also lived in a terrestrial or near-shore marine setting.” The presence of squid hooks in the stomach “does not necessarily preclude a more-normal niche in shallow water.”
‘Near shore marine’ = aquatic.
So the authors conclusion is no conclusion at all. Contra their headline, they did not ‘determine’ anything. It could have been on the beach, or in shallow water. Is the marine iguana (Amblyrhynchus cristatus) aquatic? or terrestrial? What would you say if you only found its skeleton? Or its footprints?
The good evidence continues to be
the stomach contents as the best evidence for life style (feeding niche) for the giant forms. Let the smaller ones feed in shallower waters. Let both give birth and warm up on land, like marine iguanas.
References
Bassani F 1886. Sui Fossili e sull’ età degli schisti bituminosi triasici di Besano in Lombardia. Atti della Società Italiana di Scienze Naturali 19:15–72.
Beard SR and Furrer 2017. Land or water: using taphonomic models to determine the lifestyle of the Triassic protorosaur Tanystropheus (Diapsida, Archosauromorpha). Palaeobiodiversity and Palaeoenvironments (advance online publication) DOI: https://doi.org/10.1007/s12549-017-0299-7
https://link.springer.com/article/10.1007/s12549-017-0299-7
Diedrich C 2008. Millions of reptile tracks—Early to Middle Triassic carbonate tidal flat migration bridges of Central Europe—reptile immigration into the Germanic Basin. Palaeogeography, Palaeoclimatology, Palaeoecology, 259, 410–423.
Haubold HA 1983. Archosaur evidence in the Buntsandstein (Lower Triassic). Acta Palaeontologica Polonica, 28, 123–132.
Li C 2007. A juvenile Tanystropheus sp.(Protoro sauria: Tanystropheidae) from the Middle Triassic of Guizhou, China. Vertebrata PalAsiatica 45(1): 37-42.
Meyer H von 1847–55. Die saurier des Muschelkalkes mit rücksicht auf die saurier aus Buntem Sanstein und Keuper; pp. 1-167 in Zur fauna der Vorwelt, zweite Abteilung. Frankfurt.
Nosotti S 2007. Tanystropheus longobardicus (Reptilia, Protorosauria: Reinterpretations of the anatomy based on new specimens from the Middle Triassic of Besano (Lombardy, Northern Italy). Memorie della Società Italiana di Scienze Naturali e del Museo Civico di Storia Naturale di Milano, Vol. XXXV – Fascicolo III, pp. 1-88
Peyer B 1931. Tanystropheus longobardicus Bass sp. Die Triasfauna der Tessiner Kalkalpen. Abhandlungen Schweizerische Paläontologie Gesellschaft 50:5-110.
Rieppel O, Jiang D-Y, Fraser NC, Hao W-C, Motani R, Sun Y-L & Sun ZY 2010. Tanystropheus cf. T. longobardicus from the early Late Triassic of Guizhou Province, southwestern China. Journal of Vertebrate Paleontology 30(4):1082-1089.
Wild R 1973. Die Triasfauna der Tessiner Kalkalpen XXIII. Tanystropheus longobardicus (Bassani) (Neue Ergebnisse). – Schweizerische Paläontologische Abhandlungen 95: 1-162 plus plates.
