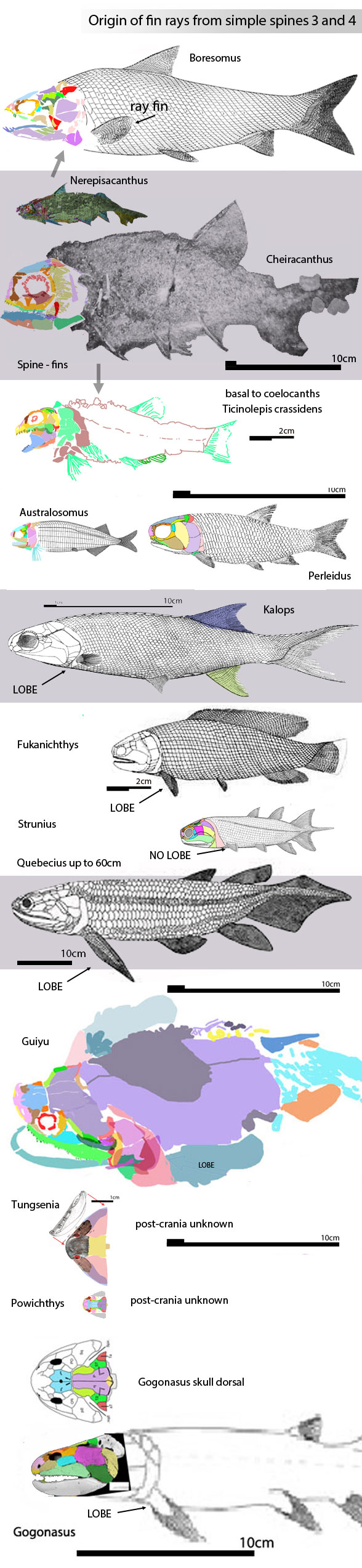
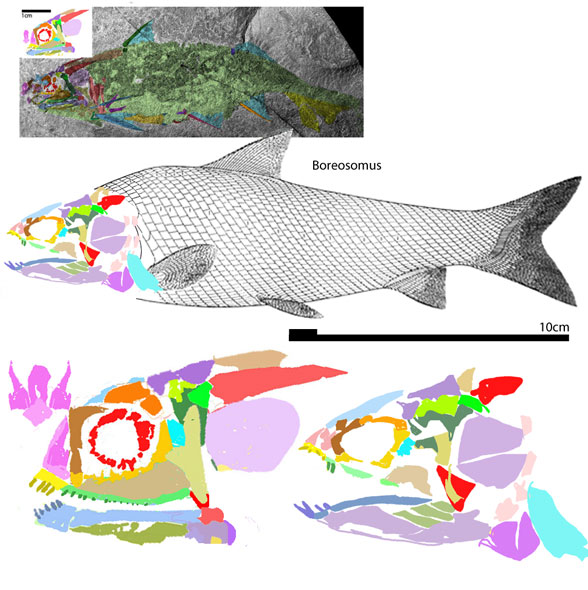
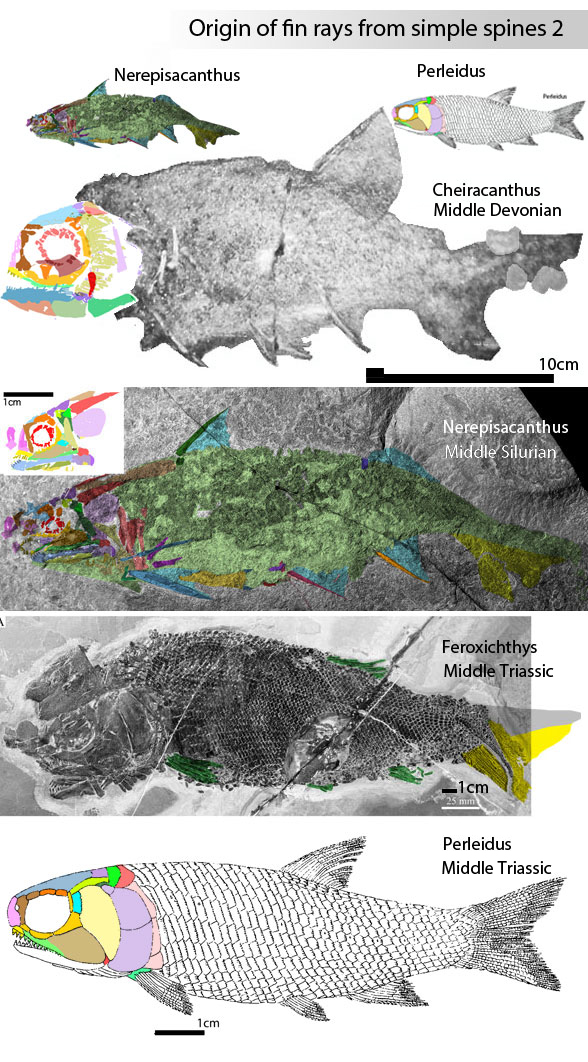
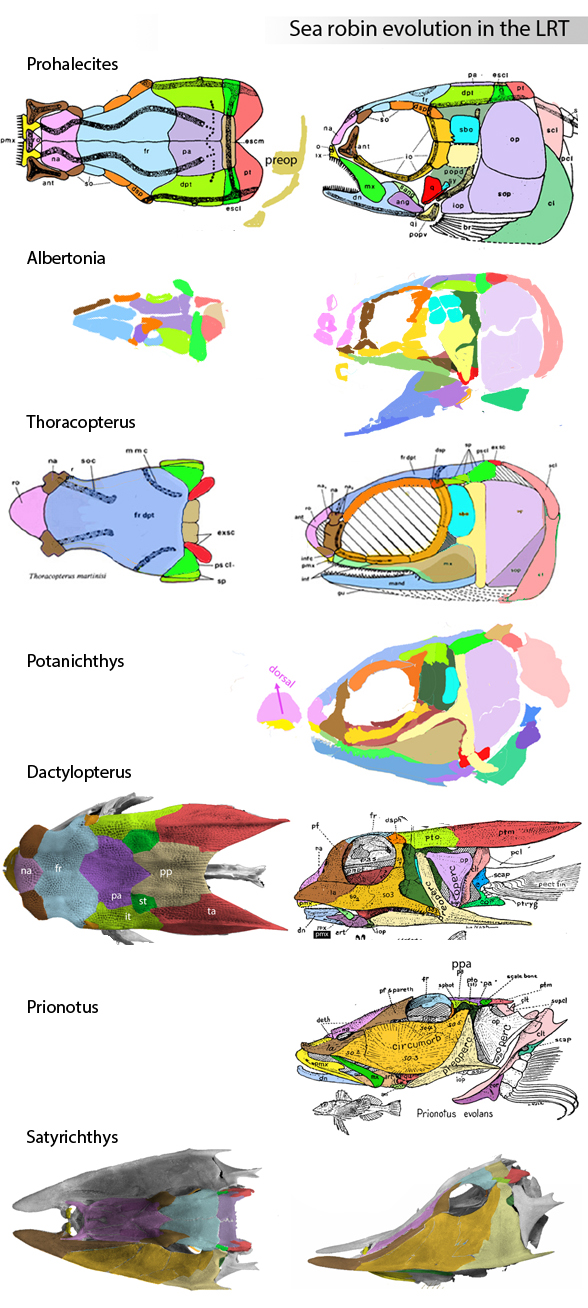
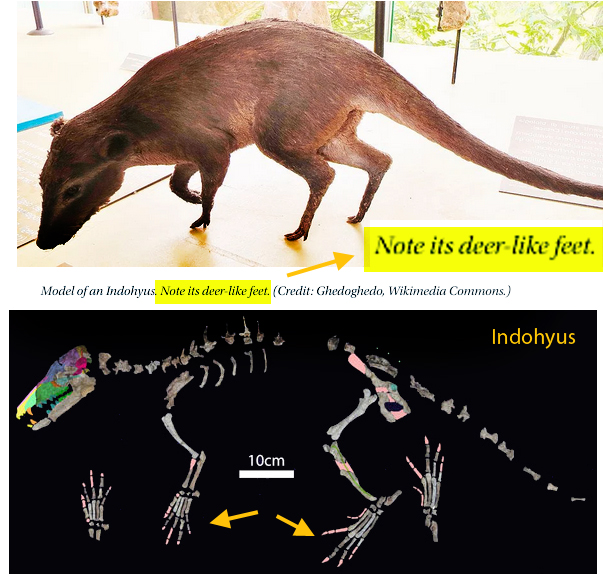
Philip Perry, writing for BigThink.com
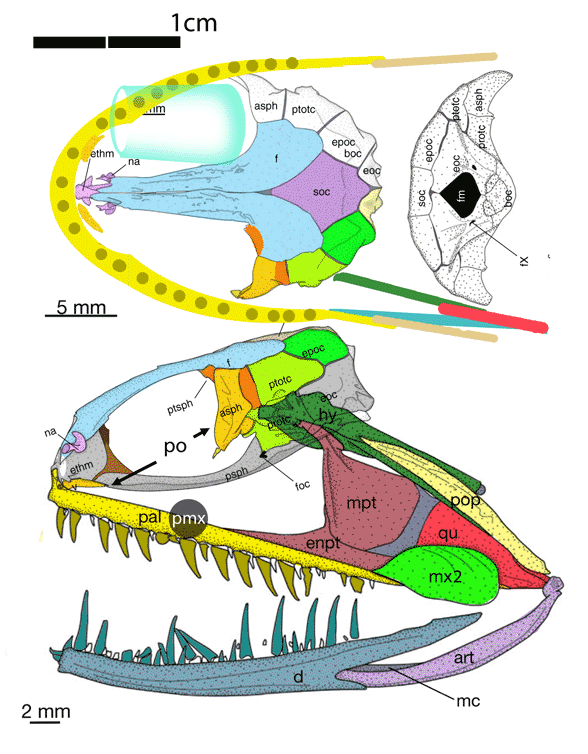
referenced a five-year-old paper by Vermeij and Motani 2018 regarding land-to-sea transitions in vertebrates. Perry’s online article was promoted with a photo of a model of Indohyus and the caption “Note it’s deer-like feet.” Unfortunately, the fossil of Indohyus does not have deer-like feet (Fig 1). As you can see, the feet of Indohyus are actually quite broad with spreading toes, like those of Pakicetus (Fig 2). Cheating anatomy is not good for science.
Promoting the deer-like hypothesis, the authors of Wikipedia wrote:
“Indohyus is an extinct genus of digitigrade even-toed ungulates known from Eocene fossils in Asia. This small chevrotain-like animal found in the Himalayas is one of the earliest known non-cetacean ancestors of whales.”
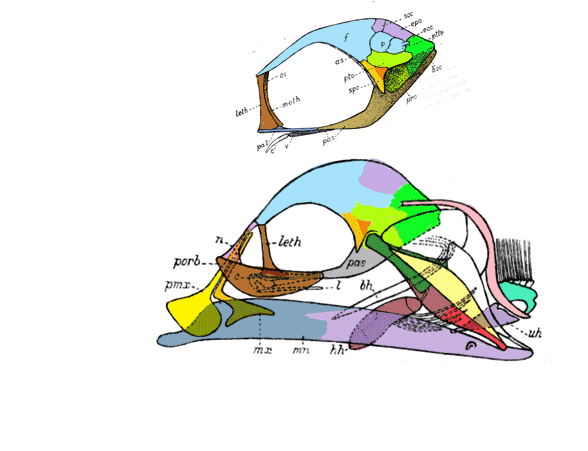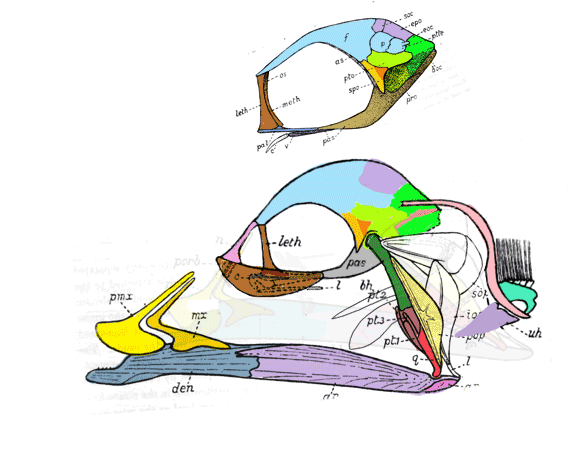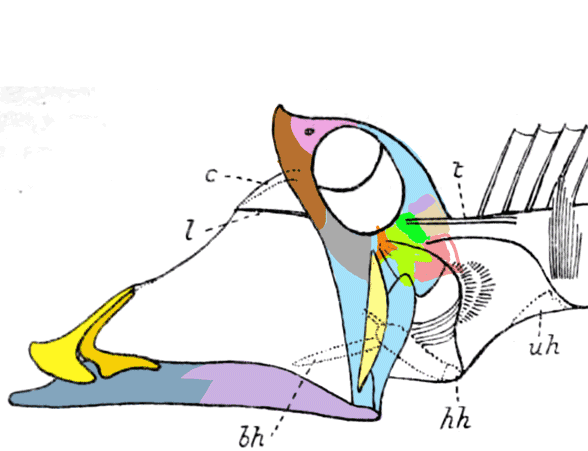
That hypothesis of interrelationships was shown to be false in 2016 due to taxon exclusion. It also does not make sense – but is widely embraced at the university level. According to the large reptile tree (LRT, 2223 taxa) Indohyus nests with an extant taxon, Tenrec (Fig 2), not with ungulates.

This all goes back to Gingerich et al 2001 and Thewissen et al 2007,
who wrote about “aquatic artiodactyls”. See figure 3.
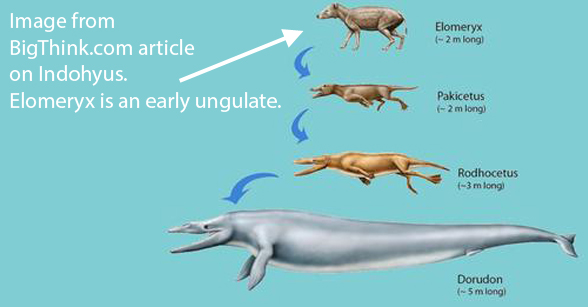
This whale origin problem could have been corrected several years ago,
but workers refereeing a manuscript submission rejected it. The manuscript points to ‘taxon exclusion’ as the only reason prior workers missed the mark. That might have been embarrassing to the academics like Gingerich and Thewiessen who mistakenly promoted the ‘early artiodactyl‘ and ‘aquatic artiodactyl’ hypothesis.
This instance of rejection was not an isolated incident, but is a common practice in paleontology. That’s why it took over a century for paleontologists to embrace the ‘birds are dinosaurs’ hypothesis and why other workers continue to embrace the ‘bat-wing bird’ hypothesis, among a slew of other bogus hypotheses presented to tuition-paying students.
You can read that rejected manuscript by Googling “Triple origin of whales”, at ResearchGate.net.
Special thanks to paleontologist, Don Prothero,
who linked the BigThink.com article to his Facebook page.
References
Gingerich, PD, Haq M, Zalmout IS, Khan IH, and Malkani MS 2001. Origin of whales from early artiodactyls: hands and feet of Eocene Protocetidae from Pakistan. Science, 293:2239-2242.
Perry P 2023. Ancient deer-like creatures returned to the ocean to become whales. But why? BigThink.com
Thewissen JGM, Cooper LN, Clementz MT, Bajpai S and Tiwari BN 2007. Whales originated from aquatic artiodactyls in the Eocene epoch of India. Nature 450:1190-1195.
Vermeij GJ and Motani R 2018. Land to sea transitions in vertebrates: the dynamics of colonization. Paleobiology 44(2):237–250. online here