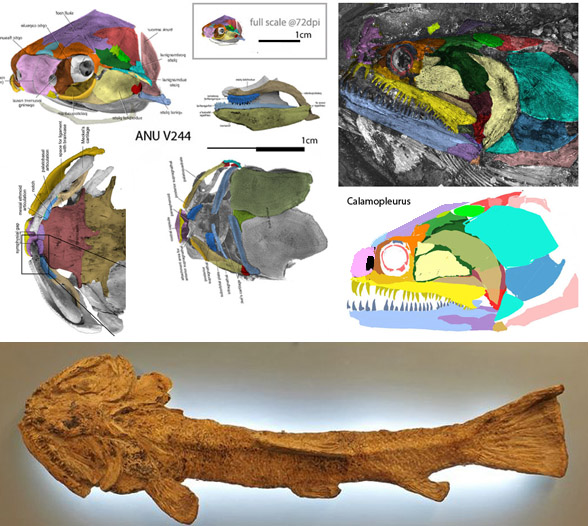
More work needs to be done on the LRT, but this current pairing of time-separated extinct fish seemed worth sharing.
Not the origin of the preopercular (Fig 1, light yellow) in this clade from a larger cheek cover in tiny toothless and unnamed ANU V244 from the Early Devonian that breaks into three parts in larger toothier Calamopleurus from the Early Cretaceous.
At the same time, note the genesis of the jugal (cyan) splitting and expanding to form a tripartite operculum in just a few hundred million years.
Figure 1. The tiny unnnamed genus (ANU V244) from the Early Devonian compared to the much larger Cretaceous predator, Calamopleurus. Note the extreme width of the skull in both along with the large coverage of the postorbital (light yellow) covering most of the cheek. The former is virtually toothless, the later is blessed with long, sharp teeth. Note also the expansion and splitting of the jugal to form a three-part operculum, convergent with several other fish clades.
References Agassiz L 1833-43. Recherches sur les poissons fossiles. Imprimerie de Petitpierre et Prince, Neuchâtel.
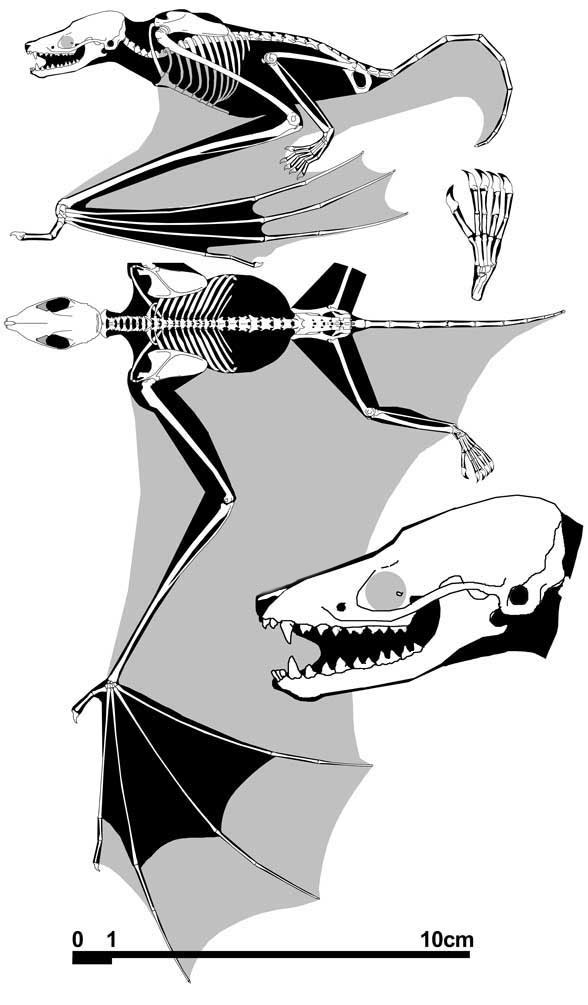
Rietbergen et al 2023 reported, “Here, we describe a new species of Icaronycteris based on two articulated skeletons discovered in the American Fossil Quarry northwest of Kemmerer, Wyoming. The relative stratigraphic position of these fossils indicates that they are the oldest bat skeletons recovered to date anywhere in the world.”
Remember: ‘oldest’ does not mean ‘most primitive.’
Figure 1. Icaronycteris gunnelli (ROM:Palaeobiology-Vertebrate Fossils:52666). One of the oldest bat fossils in the world.
We’ve seen this fossil before. I based my drawing ofIcaronycteris for ReptileEvolution.com on it in 2015 (Fig 2).
Figure 2. Icaronycteris reconstruction from several years ago based on the fossil in figure 1.
Not sure why bat experts continue to do this, but the authors of this paper cherry-picked two bat outgroup taxa not related to bats: Erinaceus, the hedgehog, and Sorex, the shrew. This is a university tradition that needs to go away.
Whale experts do this, too, nesting extinct deer, cattle and pigs basal to whales and odontocetes basal to mysticetes.
Pterosaur experts do this, too, nesting phytosaurs, Euparkeria, Lagerpeton and/or Scleromochlus as outgroup taxa… anything to avoid considering Cosesaurus.
By contrast, the large reptile tree (LRT, 2306 taxa, subset Fig 3) documents bat outgroup taxa back to Cambrian worms. The LRT nests the tiny ‘primate’ Microcebus at the base of the Chiroptera (the bat clade). Strangely, there was no mention of extant Tadarida in the text of Rietbergen et al 2023, nor does it appear in their cladogram.
BTW: The Green River bat 2022 in the LRT (Fig 3) is still unpublished.
Figure 3. Subset of the LRT focusing on bats and their ancestor, Microcebus, a mouse lemur.
Taxon exclusion continues at the end of 2023 as the number one problem facing paleontology. Never cherry-pick your outgroup taxa. Always let your wide-gamut cladogram pick for you. This is something you should be hearing from a PhD professor, not a home-schooled amateur blogger.
References Rietbergen TB, van den Hoek Ostende LW, Aase A, Jones MF, Medeiros ED, Simmons NB 2023. The oldest known bat skeletons and their implications for Eocene chiropteran diversification. PLoS ONE 18(4): e0283505. https://doi.org/10.1371/journal.pone.0283505
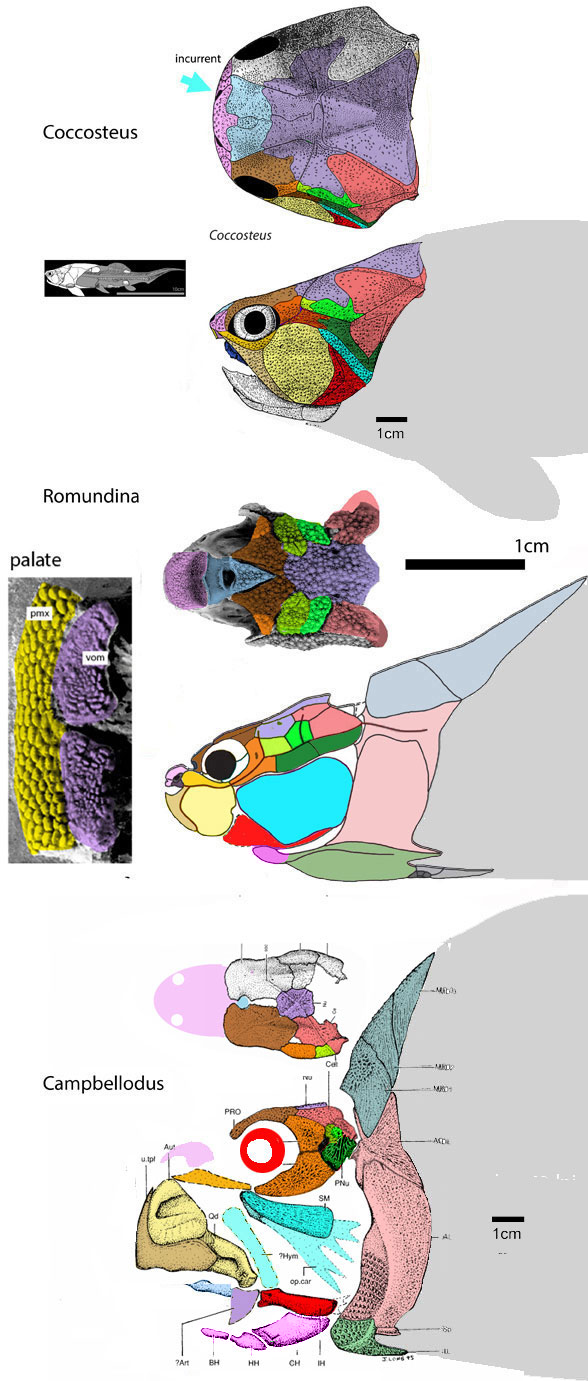
Overlooked earlier, but worth observing and considering. Let’s talk about placoderm branchials.
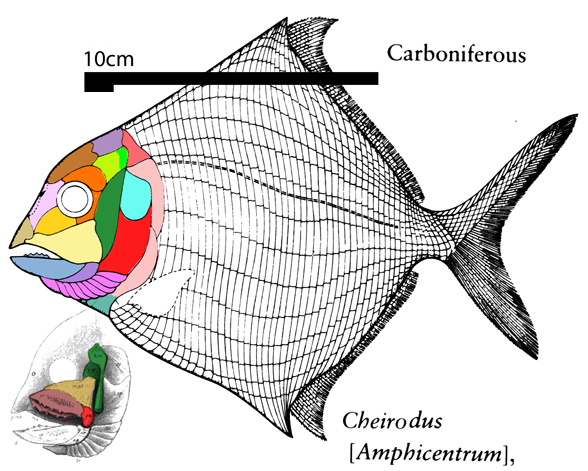
Figure 1. Coccosteus, Romundina and Campbellodus to scale. Compare to Cheirodus, Eurynotus, Bobasatrina and Platysomus below. DGS colors added here. Note the emergence of proto-branchials (magenta) in Campbellodus.
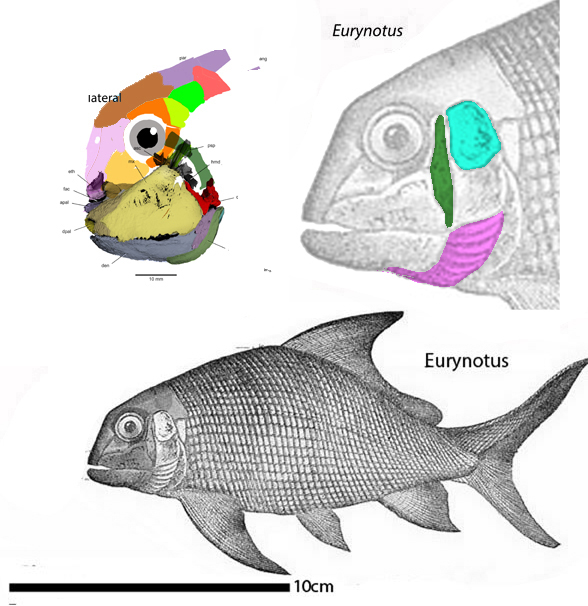
Branchials (= largely parallel strips of bone developing beneath the jaws that help expand the throat) also developed in late-surviving discoidal placoderms, like Eurynotus(Fig 2) and Cheirodus (Fig 3).
Figure 2. Eurynotus nests between Campbellodus nd Cheirodus in the LRT. Note the appearance of branchials (magenta), as in Cheirodus in figure 3.
Here we can see the initial breakup of the once solid ventral plate extending anteriorly from the plastron ofRomundina (Fig 1) and Campbellodus (Fig 1) as it splits apart forming rather typical and convergent branchials in Eurynotus (Fig 2) and Cheirodus (Fig 3).
Figure 3. Cheirodus nests as a late-surviving placoderm in the LRT. DGS colors are updated here.
Earlier Cheirodus and Eurynotus entered the placoderm clade. That hypothesis of interrelationships remains untested, unconfirmed, unrefuted and unmodified. Expand your own taxon list to test this hypothesis. Let us know what you get.
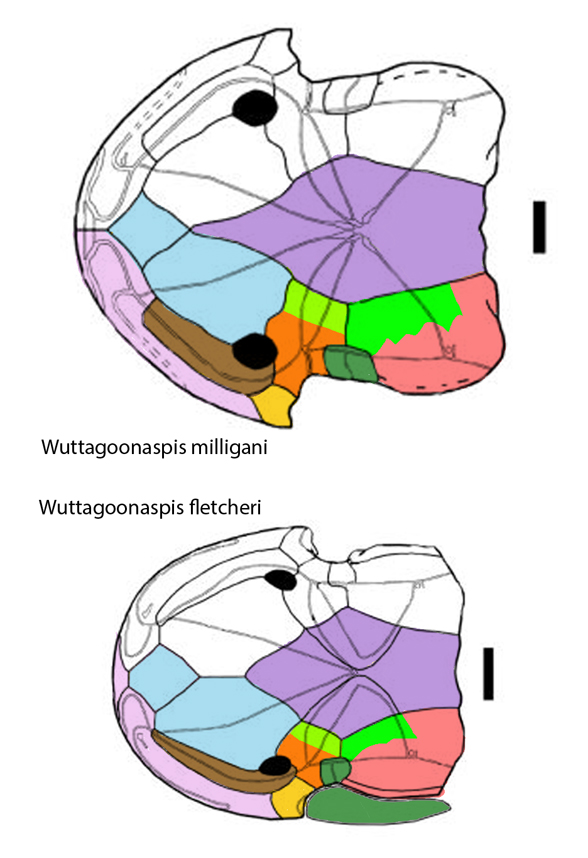
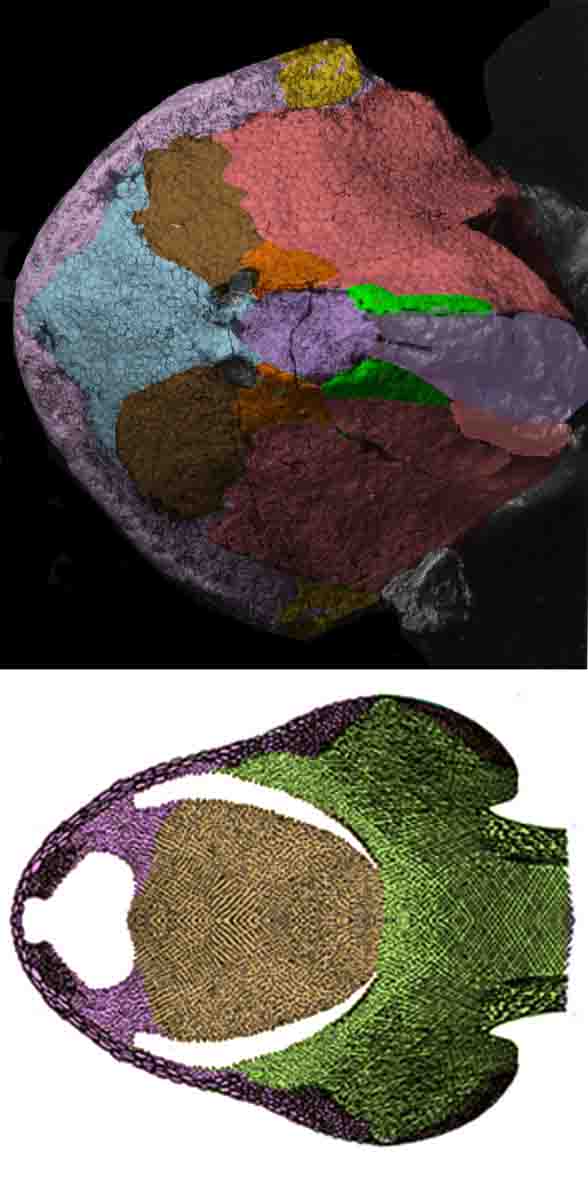
Yesterday the skull bone diagram of Middle Devonian Wuttagoonaspis(Fig 1) helped re-identify the enigmatic and largely fused skull bones of other jawless heterostracans.
Figure 1. Wuttagoonaspis. The nasals (pink) wrapp the skull perimeter.
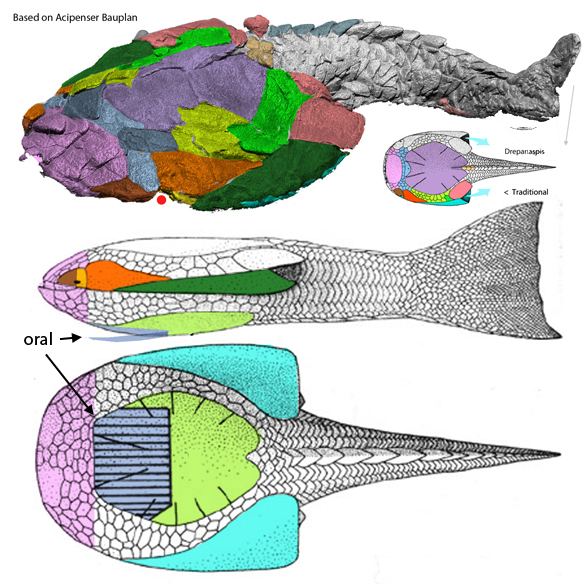
Today the same Wuttagoonaspis diagram (Fig 1) helped re-identify the enigmatic mosaic of skull bones found in sturgeons (Fig 2)… if valid.
Applying tetrapod homologs to fish skulls is done here (Figs 1, 2) to provide a common identity for scoring all vertebrates in the large reptile tree (LRT, 2306 taxa). In this way bones are free to fuse and fracture, but by giving them colors their homologs to tetrapod skull bones remain constant for scoring in the LRT.
Presently This is a hypothesis of methodology that requires confirmation, refutation or modification by other workers.
Figure 2. Acipenser skulls from Hilton 2005 and re-colored here.
You’ll note the nares of Wuttagoonaspis are ventral (not visible in figure 1), while those of the sturgeon Acipenser are lateral and confluent with the orbit. Intermediate taxa not tested in the LRT may be unknown at present. Taxon suggestions for this problem are welcome.
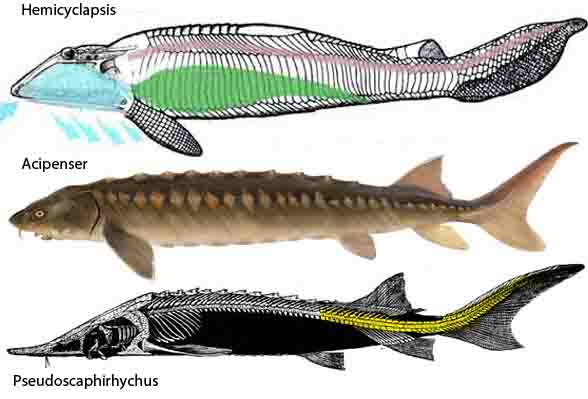
Figure 3. (Middle and bottom) two extant sturgeons, Acipenser and Pseudoscaphirhynchus, compared to the extinct heterostracan, Hemicyclaspis (top), a close relative to Wuttagoonaspis (Fig 1).
To make matters worse in the LRT more primitive taxa have terminal nares, or a single naris, located dorsally, migrating anterior to short snout from an original position between the eyes in Metaspriggina (Fig 4) in the Cambrian.
Figure 4. Metaspriggina is the most primitive taxon with a pair of eyeballs with lenses.
So the migration of the naris in basal chordates is a complex problem awaiting a solution that can be resolved with more transitional taxa that illustrate the journey of the nose around the skull in smaller, more discrete steps that those shown here (Figs 1–4).
According to Wikipedia, “Wuttagoonaspis (Fig 2) is an extinct genus of primitive arthrodire placoderm fish from the Middle Devonian of Australia.”
By contrast, in the large reptile tree (LRT, 2306 taxa) Wuttagoonaspis (Fig 2) and Kalanaspis(Fig 3) now nests basal to derived hetrerostracans, like Zenaspis(Fig 4, close to Hemicyclaspis (Fig 5). All were derived from disc-like primitive chordates like Drepanaspis (Fig 1), which has tiny lateral orbits and a ventral oral cavity.
Figure 1. Early Devonian Drepanaspis in three views. Bone colors added here.
In this hypothetical phylogenetic sequence (Figs 1–4) one can trace the migration of the lateral eyes in Drepanaspis (Fig 1) as they move step-by-step to the middle of the skull in Wuttagoonaspis and Kalanaspis (Figs 2, 3), and then to the top of the skull (Fig 4) in Zenaspis.
Figure 2. Two species of Middle Devonian Wuttagoonaspis. Colors added here.
Apparently only the front part of the skull in Kalanaspis (Fig 3) appears to be preserved in this specimen, as compared to Wuttagoonaspis (Fig 2). If instead this is a complete skull, then the posterior half has evolved away. Or the orbit is misidentified here.
Figure 3. Early Silurian Kalanaspis is the earliest known osteostracan. Here it is transitional to derived heterostracans based on the phylogenetic migration of the orbit and its nesting in the LRT.
The DGS tetrapod homology colors applied to these taxa have been revised from previous interpretations and are themselves subject to change. The Wuttagoonaspis diagram (Fig 2) provides the Bauplan for this attempt at determining where sutures might be on the suture-less skulls. Even the orbit of Kalanaspis (Fig 3 )is a best guess, revised here from prior best guesses.
Figure 4. The Early Devonian heterostracan Zenaspis documenting the final migration of the two small orbits to the top of the skull from lower locations on more primitive taxa.
Heterostracans, like Zenaspis, remain the sister clade to the extinct and extant sturgeons (Acipenseridae, Fig 5) in the LRT.
Figure 5. Two extant sturgeons, Acipenser and Pseudoscaphirhynchus, compared to the extinct heterostracan, Hemicyclaspis, a close relative to Zenaspis in figure 4. Note the similar armor.
Wuttagoonaspis fletcheri (Ritchie 1973; Middle Devonian) was originally considered ‘an unusual arthrodire’ from Australia. Here it nests basal to the heterostracan, Hemicyclaspis (Fig 5). The mouth and nares must be ventral because they are not visible dorsally.
This appears to be a novel hypothesis of interrelationships. If not, please provide a citation so I can promote it here. Housekeeping, like this, continues to improve the scoring and resolution of the fish subset of the LRT.
References Ritchie A 1973. Wuttagoonaspis gen. nov., an unusual arthrodire from the Devonian of Western New South Wales, Australia. Palaeontographica 143:58–72.
Click to view 15:25 minute video from Dr Polaris. Comments below.
Strong presentation! A few corrections based on trait analysis:
1. Madagascar elephant birds (Aepyornis maximus) are more closely related to African ostriches (Struthio). Tinamou-sized, four-toed and flightless Early Cretaceous Patagopteryx (not mentioned in the video) from Patagonia is basal to both.
2. New Zealand moas (Aepyornis) are more closely related to South American hoatzins (Opisthocomus, not mentioned in the video).
3. Kiwis (Apteryx, Fig 1) are more closely related to extant woodcock (Scolopax) and snipe (Gallinago, not mentioned in the video). Eocene Pseudocrypturus(Fig 1), a lithornithid from North America, and Late Cretaceous Vegavis, from Antarctica, are primitive members of this world-wide clade.
Figure 1. Jurapteryx, Pseudocrypturus, Apteryx and Proapteryx to scale.
4. Rheas, emus and cassowaries are giant tinamous, not as closely related to the other giant birds.
5. Genomic tests work wonders when fresh, but deliver untenable results in deep time studies that nest bats with rhinos and cats.
6. Clades should not be defined by a dozen traits, which can and do converge, but by the last common ancestor method after a wide gamut analysis of several hundred traits and taxa.
7. Be careful with your upcoming ‘notungulates’. Some traditional members are marsupials, while others are placentals. More details and cladograms at ReptileEvolution online.
Recently added to the LRT, Thaiichthys (Figs 1,2, Cavin, Deesri and Suteethorn 2013, Late Jurassic to Early Cretaceous) is used here as a skull bone/suture primer for this primitive clade of slow-moving freshwater predatory fish leading to estant gars like Lepisosteus (Fig 3). This clade is marked by subdivided skull bones with more sutures than those found in most fish skulls. That makes identity difficult.
A good Bauplan is essential. Sometimes those are hard to find.
Figure 1. Thaiichthys skeleton and scales, from Figure 2. Thaiichthys skull from Cavin, Deesri and Suteethorn 2013.
Thaiichthys buddhabutrensis is transitional between Lepidotes and Lepisosteus in the LRT. This fish is notable for its deep torso combined with a low skull resulting in a bison-like hump.
Figure 2. Thaiichthys skull from Cavin, Deesri and Suteethorn 2013. Colors added here.
The extant Cuban gar, Lepisosteus (Fig 3), really needs a good Bauplan, like Thaiichthys (Fig 2), to identify the plethora of subdivided skull bones, an essential first step prior to scoring for analysis. The diagram and written labels reflecting traditional identities are from Gregory 1933.
Figure 3. Lepisosteus, the extant Cuban gar, with bones newly colorized with Thaiichthys homologies. The yellow-green and green colors on the dorsal cranium indicate the location of the former intertemporal and supratemporal respectively.
Nothing here can be accomplished without earlier discoveries, studies and publications. When you attempt a phylogenetic analysis of fish be prepared for the unexpected. A correction made in one clade will sometimes ripple through the entire tree. You will learn something in the process: Patience. BTW, I should not be the only worker attempting this.
References Cavin L, Deesri U and Suteethorn V 2013. Osteology and relationships of Thaiichthys nov. gen:A ginglymodi fom the Late Jurassic – Early Cretaceous of Thailand. Palaeontology 56(1):183–208. Gregory WK 1933. Fish skulls. A study of the evolution of natural mechanisms. American Philosophical Society 23(2) 1–481.
Here is a case where the traditional diagram does not match the available in situ photograph (Fig 1). Previously I scored Miguashaia from the diagram. Now scores are based on the photograph of the fossil.
Miguashaia bureaui (Schultze 1973, Cloutier 1996; Middle Devonian; 45cm) was considered the sister group (outgroup) of the Actinista (coelocanths). Notably Miguashaia retains a heterocercal tail and other sarcopterygian and osteichthyan traits. The maxilla was considered taphonomically missing, but the premaxilla is wide. The dentary in the reconstruction is a standard coelacanth design, not what we see in the fossil in situ. Compare to Late Silurian Guiyu, with which it nests in the LRT. Perhaps another specimen more closely matches the diagram shown here.
Miguashaia bureaui (Schultze 1973, Cloutier 1996; Middle Devonian; 45cm) was considered the sister group (outgroup) of the Actinista (coelocanths). Notably Miguashaia retains a heterocercal tail and other sarcopterygian and osteichthyan traits. The maxilla was considered taphonomically missing, but the premaxilla is wide. The dentary in the reconstruction is a standard coelacanth design, not what we see in the fossil in situ. Compare to Late Silurian Guiyu, with which it nests in the LRT. Note the wide flat skull and torso vs the tall narrow hypocercal tail.
It is only a coincidence that a similar ‘bite mark’ is found in the dorsal surface of both flattened taxa (Figs 1,2).
Figure 2. Late Silurian Guiyu with catfish homologs. See Miguashaia in figure 1 for comparison. The spine is a former placoderm carapace, now reduced and elevated.
Guiyu oneiros (Zhu et al. 2009; late Silurian; V17914) was originally considered the oldest osteichthyan and a member of the lobe-fin clade close to Psarolepis. This revised restoration gives this taxon almost a sea-turtle like appearance, with large pectoral paddles directed posteriorly. Guiyu is transitional from placoderms, but closer to catfish in the LRT. The nose and chin are unchanged inthe catfish, Clarias.
Still struggling here with the fish subset of the LRT. I’m looking at every one of the 490+ taxa, making corrections where appropriate. Example shown above.
References Cloutier R 1996. The primitive actinistian Miguashaia bureaui Schultze (Sarcopterygii). Pp. 227–247 in: Schultze H-P and Cloutier R editors. Devonian fishes and plants of Miguasha, Quebec, Canada. Dr. Friedrich Pfiel; München: 1996. Schultze H-P 1973. Crossopterygier mi heterozerker Schwanzfloss aus dem Oberdevon Kanadas, nebst einer Beschreibung von Onychodontida-Resten aus dem Middledevon Spaniens und aus dem Karbon der USA. Palaeontograhica A 143:188–208. Zhu M, Yu X-B and Janvier P 2014. A primitive fossil fish sheds light on the origin of bony fishes. Nature 287:607–610. Zhu M, Zhao W, Jia Lu J, Qiao T and Qu Q 2009. The oldest articulated osteichthyan reveals mosaic gnathostome characters. Nature 458:469-474.
Once again, this solution became obvious when the problem was finally solved. These two taxa (Fig 1) were never far apart, but they finally came together in the LRT.
Figure 1. Monocentris compared to its tiny Cretaceous relative, Cretatriacanthus. This connection might have come sooner if I had imagine erect dorsay spines on Monocentris, here confirmed with an invivo photo. Here again scale bars are important for showing phylogenetic miniaturization at the genesis of new morphologies. Note: the pelvic fins are here transformed into ventral spines, as in spiny sharks and sticklebacks.
Monocentris japonicus (originally Scomber japonicus Houttuyn 1782; 12-17cm) is the extant Japanese pineconefish, so named because it is covered in scutes, a larger sort of scale. This nocurnal fish lives in caves. It has light-producing areas filled with bioluminescent bacteria on the lower jaws, distinct from the unrelated flashlight fish, Anomalops. Due to the restrictive armor, propulsion comes from the pectoral fins, as in the related taxa, Lampris and Exocoetus.
Cretatriacanthus guidottii (Sorbini 1996, Fig. 1; early Late Cretaceous; about 3cm) is traditionally known as the earliest known tetraodontiform, the clade that includes queen trigger fish, ocean sunfish and pufferfish. Here it nests with equally short moonfish like Mene and much longer oarfish, like Regalecus along with its newest relative, Monocentris (Fig 1).
References Houttuyn M 1782. Beschryving van eenige Japanese visschen, en andere zee-schepzelen. Verh. Holl. Maatsch. Wet. Haarlem. 20(2): 311-350. Sorbini L 1979. Segnalazione di un plettognato Cretacico Plectocretacicus nov. gen. Bollettino del Museo Civico di Storia Naturale di Verona, 6:1–4.
Earlier the large reptile tree (LRT, 2306 taxa) recovered three separate origins for the pectoral girdle and pectoral fins. Here’s one that popped up recently, the one that ultimately led to tetrapods, like us.
Figure 1. Coilia is traditionally considered a type of herring (Clupeiformes), but here shares traits with Silurian bony fish, like Lasanius, that first developed pectoral fins. Note the enlarged rays on both taxa and that odd nose they both share, distinct from other fish. We also see one origin of the anal fin and dorsal fin here.
430 million years separate Middle Silurian Lasanius frrom extant Coilia, the deep-sea grenadier anchovy. Yet both share a suite of traits that bring these two together in the LRT.
Figure 2. The skulls of Lasanius and Coilia compared. Note the long maxilla (green) in both taxa, hyperelongated in Coilia.
The European anchovy, Engraulis, is a more plesiomorphic ancestor for tetrapod ancestors. Both extant taxa have been evolving for hundreds of millions of years, so teasing out these interrelationships has been difficult. The other two origins of fins and opercula, in sturgeons and placoderms, were remarkably similar in all respects, except their genesis. This hypothesis of interrelationships requires confirmation, refutation or modification.