In order to know taxa better
it’s worthwhile to revisit earlier interpretations once a bit of experience working with clade patterns is gained. The is the next step beyond freshman naiveté. In this case, several taxa (Figs 1–4) were revisited after earlier interpretations beset with first-timer mistakes. The resulting new interpretations update and correct prior errors and improve analytical scores.

Early Silurian Shenacanthus
(Fig 1, Zhu et al 2022) has a 5mm skull crushed and somewhat scattered. Here, after a bit of experience and sweat equity, Shenacanthus is re-reconstructed. Now it is even more similar to the much larger, much later big-eyed basal shark, Ozarcus.
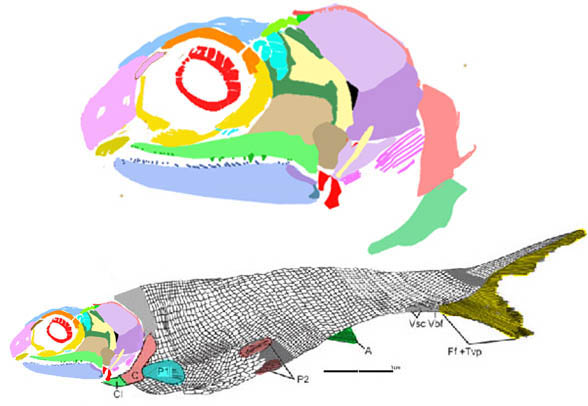
Early Carboniferous Ozarcus
(Fig 1, Pradel et al 2014) was originally µCT scanned, so it is perfectly presented. Earlier I mistook the strip-like palatine for a maxilla. This simple correction was made by changing its DGS color to cyan. Now no chondriichthyans have a maxilla.
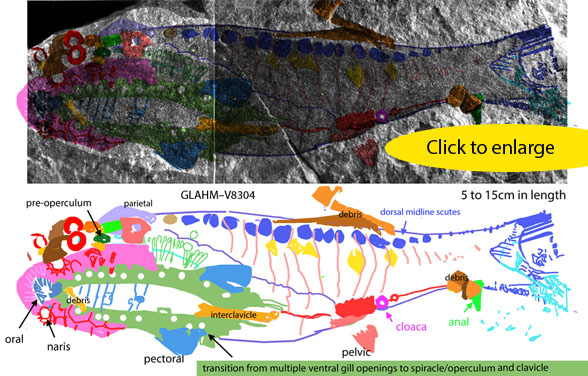
Silurian Thelodus
(Fig 2) is a dark on dark 2D fossil that was difficult to sort out and interpret. Here, after some experience and several stumbling attempts DGS colors now indicate this specimen was basal to sturgeons with small fins and ventral nares. This linking Thelodus to heterostracans, and indicates their progress in their migration to their eventual position anterior to the orbits in extant sturgeons. A tiny hyomandibular disc (dark green) makes an appearance near the spiracle, prior to its enlargement as yet another type of operculum. FInally, if not yet another piece of debris, the interclavicle makes an appearance at the flat ventral midline of Thelodus. This bone is retained in later placoderms, which underlap it with lateral elements.

Devonian Euphanerops,
(Fig 3) is a lamprey-like, lancelet-like (Fig 4) relative of Bikenia and the conodont, Promissum in the LRT. Like conodonts, Euphanerops had ‘jaws’ made up of several separate slender elements which linked more tightly together in more derived gnathostomes basal to tetrapods. Here the maxillae form an arc without a premaxilla. By contrast, the early dentary elements were straight. Conodonts can have a dozen of these, but Euphanerops has fewer. Here all the jaw elements lack teeth or conodont-like sharp processes.

The pelvic fins of Devonian Euphanerops,
(Fig 4, Woodard 1900; Janvier and Arsenault 2007) were identified as unusual paired anal fins by Sansom, Gabbott and Purnell 2013 because the atrium excurrent valve was miisidentified as the anus (= the cloaca). Here corrections are offered in DGS colors. So far I have not seen any paired anal fins.

All the fins on the Carboniferous eel-like Paratarrasius
How would you score this taxon? Did it have a straight tail? Or a hetercercal tail? Comparie Paratarrasius (Fig 5) to its LRT sister, Bluefieldius (Fig 6), which has a heterocercal tail. That makes the dorsal fin of Paratarrsius extend to the tail tip (not beyond as the diagram (Fig 5, lower right) shows. In addition, pelvic and anal fins were originally overlooked based on the finless diagram. The pectoral fins were imagined with dashed lines to look like ping-pong paddles. Here all laferal fins are vestigial, but still present. They are given DGS colors for identification (Fig 5) and appropriate scores in the LRT.
These corrections
are improving the scoring of the fish subset of the LRT, which is still ‘in progress’. Perhaps it will always be ‘in progress.’
References
Janvier P and Arsenault M 2007. The anatomy of Euphanerops longaevus Woodward, 1900, an anaspid-like jawless vertebrate from the Upper Devonian of Miguasha, Quebec, Canada. Geodiversitas 29 (1) : 143-216.
Pradel A, Maisey JG, Tafforeau P, Mapes RH and Mallant J 2014. A Palaeozoic shark with osteichthyan-like branchial arches. Nature 13185. doi:10.1038/nature13195e
Sansom RS, Gabbott SE and Purnell MA 2013. Unusual anal fin in a Devonian jawless vertebrate reveals complex origins of paired appendages. Biol Lett. 2013 Jun 23; 9(3): 20130002. doi: 10.1098/rsbl.2013.0002
Zhu Y-A et al (10 co-authors) 2022. The oldest complete jawed vertebrates from the early Silurian of China. Nature 609:954–958. online
Woodward AS 1900. On a new ostracoderm fish (Euphanerops longaevus) from the Upper Devonian of Scaumenac Bay, Quebec, Canada. Magazine of Natural History ser. 7, 5: 416-419.
wiki/Tarrasiiformes
wiki/Thelodus
wiki/Euphaneropidae
wiki/Shenacanthus – not yet posted