
Sorry this took so long…
As you’ll see there was a lot of work and prep involved that has been several weeks in the making. Thank you for your patience.
Earlier I introduced the Marjanovic and Laurin 2016 study
the way they did, by reporting their confirmation of the Ruta and Coats 2007 basal tetrapod topology that they were testing prior to reevaluating the data. I noted then that both studies (Fig. 5) included many so-called pre-reptiles, including Bruktererpeton, Chroniosaurus, Solenodonsaurus, Limnoscelis, Tseajaia, Diadectes, Orobates and Westlothiana,should not be in the pre-amniote inclusion set. Those taxa nest within the Reptilia in the large reptile tree (LRT, subset Fig. 4) with Silvanerpeton and Gephyrostegus at the base of the Reptilia (= Amniota). As reported earlier, those two are the amphibian-like reptiles that first developed the amniotic egg that defines the clade Amniota, a junior synonym of the Reptilia, based on the tree that recovers them at the base of both major branches, the new Archosauromorpha and the new Lepidosauromorpha early in the Viséan.
How can one readily compare two competing cladograms?
You would not want to sit through a comparison of tens of thousands of scores for competing trees in a short blog like this. But we can compare images of taxa (Figs. 1–3. 6–8) placed in their phylogenetic order, subdivided for clarity into the three major lineages of basal tetrapods:
- Basalmost tetrapods and the lineage that led to Reptilia
- Members of the Lepospondyli
- Members of the Microsauria
These images will serve as a ready reference for today’s topics. As a preview, in summary:
The Marjanovic and Laurin (ML) 2016 tree nests
- frogs like Rana and salamanders like Andrias with microsaurs.
- small amphibamids, Cacops and Micromelerpeton nest with temnospondyls.
- basal Amniota splits into Synapsida (Caseasauria + Archaeovenator) and Sauropsida (Captorhinus, Paleothyris, Petrolacaosaurus) arising from an unknown genus basal to Diadectomorpha + Amniota
- The clade Amphibia arises near Solenodonsaurus + the crown-group Tetrapoda
- The clade Microsauria is divided into three parts separated by non-microsaurs with origins near Westlothiana.
The LRT nests
- frogs and salamanders nest with lepospondyls.
- small amphibamids, Cacops and Micromelerpeton nest with lepospondyls.
- basal Amniota splits into Archosauromorpha (several basal taxa, Archaeovenator, Paleothyris and Petrolacaosaurus) and Lepiodosauromorpha (several basal taxa, Caseasauria and Captorhinus) with both major clades arising from Gephyrostegus bohemicus a late-surving Westphalian taxon, and Silvanerpeton, a Viséan taxon.
- The clade Amphibia arises near Balanerpeton and the amphibamids.
- The clade Microsauria has a single origin near Kirktonecta
What you should be looking for
is a gradual accumulation of traits in every lineage. And look for taxa that don’t fit in the order presented. This can be done visually with these figures, combining hundreds of traits into one small package. Rest assured that all scoring by ML and the competing analysis in the LRT were done with the utmost care and diligence. So, some biased or errant scoring must have taken place in one study or the other or both for the topologies to differ so great. Bear in mind that ML had firsthand access to fossils and may have bowed to academic tradition, while I had photos and figures to work with and no allegiance to academic tradition.
First
the large reptile tree (LRT) taxa (Figs. 1–3) had two separate origins for limbed vertebrates.

Figure 1. CLICK TO ENLARGE. Basal tetrapod subset according to the LRT. These taxa lead to Reptilia, Lepospondyli and through that clade, the Microsauria. Note the convergent development of limbs and digits arising out of Osteolepis.
In both studies
basal tetrapod outgroups are tail-propelled sarcopterygians having muscular fins not yet evolved into limbs with digits. Behind the skull are opercular bones that are lost in taxa with limbs. An exoskeleton of bony scales disappears in taxa with limbs. Snout to tail tip length averages 50 cm.
In the LRT
locomotion switches to the limbs in temnospondyls, which tend to be larger (1m+ and have overlapping dorsal ribs. The Greererpeton branch flattens out the ribs and skull, reducing both the tail and the limbs to likely become sit-and-wait predators. Phylogenetic size reduction and limb elongation is the trend that leads to Reptilia (Gephyrostegus). However an early exception, Crassigyrinus (Fig. 1), elongates the torso and reduces the limbs to adopt an eel-like lifestyle. Kotlassia adopts a salamander-like lifestyle from which Utegenia and the Lepospondyli arise (Fig. 2) alongside Reptilia.

Figure 2. CLICK TO ENLARGE. Subset of the LRT representing lepospondyli leading to frogs.
In the LRT,
short-tailed, salamander-like Utegenia (derived from the Seymouriamorpha, Fig. 2) is a late-surving basal member of the generally small-sized clade Lepospondyli, which ultimately produces salamanders and frogs. A side branch produces the larger, temnospondyl-like Cacops, which develops a bony ridge atop the dorsal spines. Note the nesting here of Gerobatrachus as a salamander and frog relative, distinct from the ML tree (Fig. 6).

Figure 3. CLICK TO ENLARGE. Subset of the LRT focusing on Microsauria.
In the LRT
the Microsauria are derived here from the small basal amphibamids, Caerorhachis and more proximally, Kirktonecta. Microsaurs range from salamander-like to lizard-like to worm-like. The tail elongates to become the organ of locomotion in the Ptyonius clade. The head and torso flatten in the Eoserpeton clade.
Below
is the pertinent subset of the LRT (Fig. 4) with a representative, but not complete or exhaustive set of taxa. A summary of the tree’s differences with the ML tree is presented above. The ML tree is summarized below in three parts (6-8).

Figure 4. Subset of the LRT focusing on basal tetrapods.
The Marjanovic and Laurin 2016 tree
(Fig. 5) presents a topology that is similar to the LRT in parts, but distinct in other parts, as summarized above. I realize this presentation is illegible at this column size due to the large number of taxa. Click on it to enlarge it. At the top and down the right column are basal taxa leading to temnspondyls and reptiles at bottom right. Working from the bottom up the left side are the microsaurs ending with the lissamphibians (frogs and salamanders) at the top/middle of the left column.
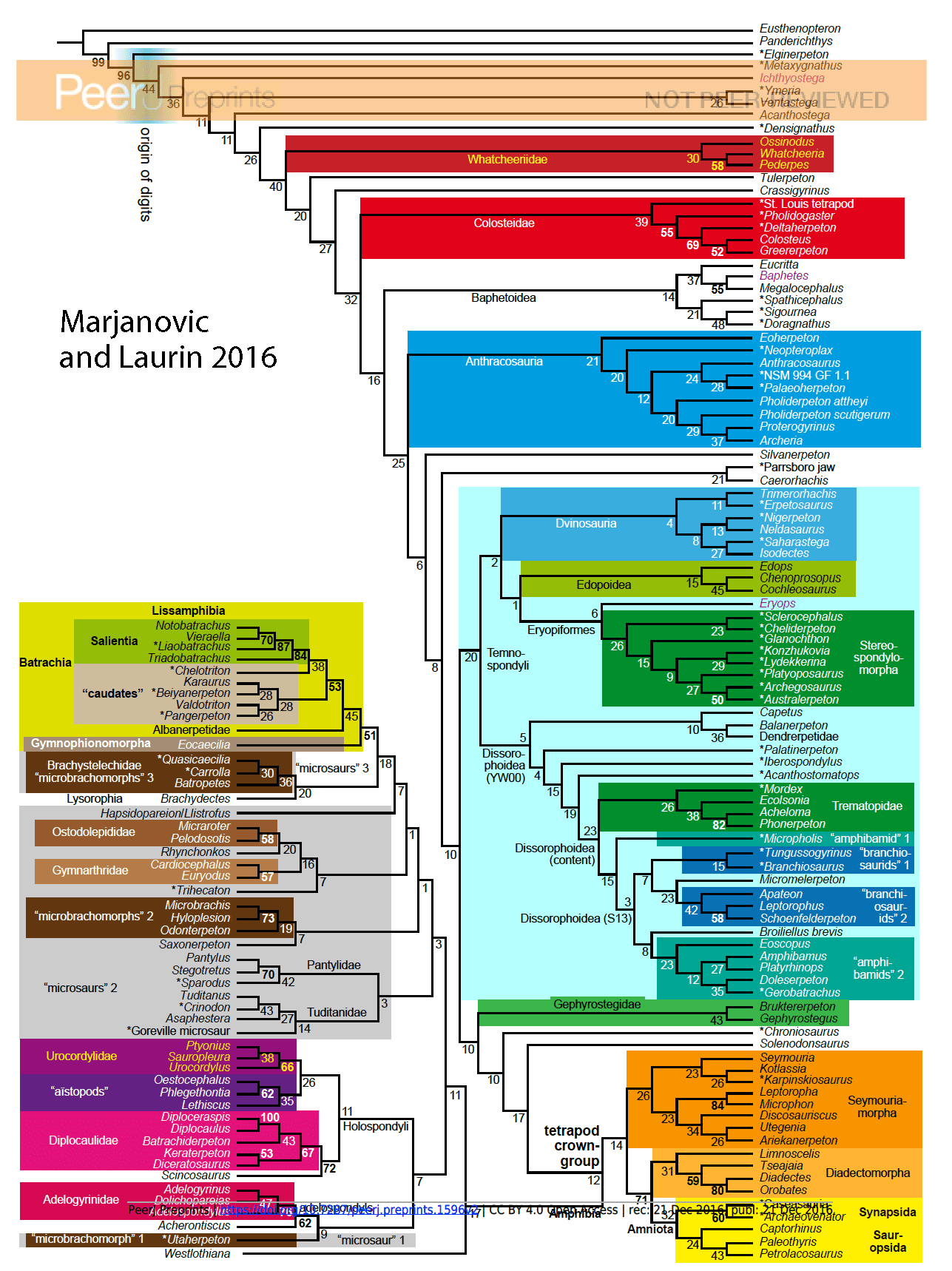
Figure 5. CLICK TO ENLARGE. The reevaluated Marjanovic and Laurin tree from which taxa on hand were set to match the tree topology (Figs. 5-7).
The ML tree
subdivides into there parts (Figs 6-8): basal taxa, some leading to temnospondyls and amphibamids; taxa leading to and including Amniota; and finally microsaurs leading to and including extant amphibians.

Figure 6. Basal tetrapods according to Marjanovic and Laurin 2016. Figures 6 and 7 lead to Amniota and Microsauria respectively.
In the ML topology,
Ichthyostega, a taxon with a very large pectoral girdle, ribs, and pelvis, gives rise the the altogether smaller and more fish-like Acanthostega, which gives rise to members of the Whatcheeridae, tall-skulled Crassigyrinus and flat-skulled Osinodus. The traditional Colosteidae arise next. They have a variety of long shapes with short-legs. Oddly from this seemingly primitive clade arises small, short-torsoed, long-legged Eucritta followed by long torsoed, short-legged Proterogyrinus followed by a large clade of short-torsoed, long-legged taxa, including the >1m temnospondyls and the <30cm amphibamids.

Figure 7. CLICK TO ENLARGE. These are taxa listed on the Marjanovic and Laurin 2016 that lead to Reptilia (Amniota).
In the ML tree
Gephyrostegus arises from the small temnospondyl, Balanerpeton, and and gives rise to Chroniosaurus, Solenodonsaurus, the Seymouriamorpha (including Utegenia) and the Diadectomorpha, nesting as the sister clade to the Amniota. Thus, no phylogenetic miniaturization was present at the origin of the Amniota in the ML tree. Moreover, dozens of taxa were not included here that nest at the base of the Amniota (Reptilia) in the LRT. Basal amniotes in the ML tree are all Latest Carboniferous to Early Permian, while in the LRT basal amniotes arrived at least 40 million years earlier in the Visean (Early Carboniferous) and had radiated widely by the Late Carboniferous, as shown by the ML taxaon list. No amphibian-like reptiles made it to their Amniota.

FIgure 8. CLICK TO ENLARGE. Microsauria according to Marjanovic and Laurin 2016. Here frogs and caecilians nest within the Microsauria.
In the ML tree
the three microsaur clades (Fig. 5) arise from the Viséan taxon, Westlothiana (Fig. 8), which nests as a derived reptile when tested against more amniotes in the LRT. Utaherpeton is a basal microsaur in both trees, but it gives rise to the eel-like Acherontiscus and kin in the ML tree. Westlothiana further gives rise to Scincosaurus and kin, including the larger Diplocaulus. Thirdly, Westlothiana gives rise to lizard-like Tuditanus which gives rise to big-skulled Pantylus and tiny-limbed Microbrachis, shark-nosed Micraroter and Rhynchonkos. In both trees, Batropetes bucks the long-body, short-leg trend. In both trees Celtedens, representing the salamander-like albanerpetontids, gives rise to extant salamanders and frogs
So the possibilities are:
- Only one tree is completely correct
- Only one tree is mostly correct.
- Both trees have some correct and incorrect relationships
Problems
- Basal tetrapods tend to converge on several traits. For instance in the LRT, the palate is ‘open’ with narrow pterygoids in both temnospondyls and lepospondyls.
- Many small derived taxa lose and fuse skull bones
- Many taxa fuse vertebral bones as they evolve away from the notochord-based semi-encircling vertebrae of fish toward more complete vertebrae in which the neural spine, pleurocentrum and intercentrum tend to fuse, sometimes in convergent pattern, as widely recognized in basal reptiles and microsaurs.
- In basal tetrapods, fingers are not often preserved. So when four fingers appear their identity has to be ascertained. In the LRT mc5 and digit 5 are absent in Lepospondyls. In the LRT mc1 and digit 1 are absent in the temnospondyls. Five fingers and/or metacarpals are preserved in the few other non-amniote, basal tetrapods that preserve fingers (Proterogyrinus, Seymouria). The ML tree assumes that when four digits are present, they represent digits 1–4.
Ultimately
maximum parsimony and Occam’s Razor should rule unless strong evidence to the contrary is provided. After evidence is presented, it’s up to colleagues to accept or reject or ignore hypotheses.
References
Marjanovic D and Laurin M 2016. Reevaluation of the largest published morphological data matrix for phylogenetic analysis of Paleozoic limbed vertebrates. PeerJ. Not peer-reviewed. 356 pp.
Ruta M and Coates MI 2007. Dates, nodes and character conflict: addressing the lissamphibian origin problem. Journal of Systematic Palaeontology 5-69-122.

…I know you’re not doing that deliberately, but apart from that I feel as if I had just witnessed a Gish gallop. It’s sad that you still haven’t begun to read the preprint; you’ve only looked at a different picture than last time.
Particularly baffling is the fact that you didn’t read the comments to your previous post on this preprint. One of the comments points out that you cited the preprint wrong. You still cite it wrong. PeerJ PrePrints is not PeerJ, and the preprint has an article number and even a volume number. (…And, BTW, can’t you copy & paste the ć at the end of my name? The accent isn’t ornamental, it changes the pronunciation pretty drastically.)
First of all, we didn’t produce a single tree. We made twelve parsimony analyses, four of them unconstrained – and your fig. 5 presents none of them! It shows one of our four bootstrap trees. Do you know how bootstrapping works, and why it routinely gives you trees that are different from all of the most parsimonious trees?
Second, it is called sister-group, not mother-group, for a reason! It really blows my mind that this has never occurred to you in the last twelve or more years. Sister-groups have a common ancestor that was different from both of them, indeed from all of the taxa in the whole data matrix, except in highly exceptional cases. None of the leaves of our trees is an ancestor of any other. This is normal.
BTW, if anyone is looking for Cacops in any of our trees, it’s not there; the closest we have is its fellow dissorophid Broiliellus brevis (which may or may not end up belonging to Broiliellus once the Broilellus–Dissorophus mess is sorted out).
Uh, Microsauria is a (paraphyletic) subset of Lepospondyli, and your usage of the term “lineage” is quite unlike any in the literature. How about:
1. Stem-tetrapods and the amniote total group
2. Amphibians rootward of the “microsaur” grade (in one word, Westlothiana)
3. The clade of amphibians other than Westlothiana
Actually, in those analyses that include any synapsids at all, we find Diadectomorpha inside Amniota, on the synapsid side.
Let’s call the whole synapsid side Theropsida Goodrich, 1916; in that case, Theropsida and Sauropsida are sister-groups, and they both arise from the first amniote. Y’know, good old Bob the Basal Amniote.
Genera don’t exist outside of taxonomists’ heads.
By definition, the tetrapod crown-group consists of two branches and their last common ancestor; and also by definition, one of these two branches is called Amphibia – it consists of everything that’s more closely related to Lissamphibia than to Amniota.
(Lissamphibia is of course more closely related to itself than to Amniota, so it’s a subset of Amphibia.)
Specifically, we find “Microsauria” not to be a clade, but a paraphyletic assemblage; and we find the smallest clade that contains all “microsaurs” to be the sister-group of Westlothiana.
No, I’m afraid. No. Neither the coding (making the characters) nor the scoring (filling in the cells in the matrix) for the LRT were done with the utmost care or diligence. In fact, as I’ve already demonstrated in comments to an earlier post, it was done with blissful ignorance of anatomy, phylogenetics, development biology, practices of publication and illustration (like omitting scales from drawings), and probably a few other things.
Or, of course, the character samples are different. Guess what! They’re extremely different! There’s a huge difference between our matrices right there!
Odd how you don’t mention them with one word. It would be easy to criticize the character sample in the preprint – in fact, I’ve been adding characters to plug the most glaring holes in it since a few weeks ago, and I’ll keep going for the next three years if the grant proposal for that is accepted. But no, you’re letting this obvious opportunity zoom past your head. Why is that?
Nowhere near as many as we’d have liked to.
Ah, you don’t let people cuss, but you insult them freely? Show us some evidence for your claim that we haven’t thought for ourselves.
Oops, I mistyped the </blockquote> at the end of your claim about scales.
It seems to have taken me about an hour to write this comment. It’s 1 am now, I’m off to bed.
Oh, another thing: your reconstructions of the body outline of frogs, salamanders and amphibamids are way off. You’d know that if you’d looked at extant frogs, salamanders or caecilians or even at fossil frogs and salamanders with preserved body outlines. The ribs of all these animals are extremely short; they don’t reach the side wall of the body, but end long before it.
That’s how caecilians manage to move the vertebral column + ribs + inner muscle layer independently of the skin + outer muscle layer.
Bear in mind that ML had firsthand access to fossils and may have bowed to academic tradition
Are you kidding me? Do you have any knowledge at all of the broader trends in orthodox vs unorthodox hypotheses of tetrapod phyogeny? Do you have any understanding of hat Marjanovic & Laurin are even doing in this paper? Do you even understand what the big debates are in early tetrapod evolution?
The trees that M&L present are far from orthodox, and neither of them would say that this is the final word on early tetrapod phylogeny. M&L are specifically testing whether existing analyses contain strong support for any specific relationships of lissamphibians, and what the effects are of changing character delineation techniques. Essentially, they are asking whether the disagreements between working groups on lissamphibian origins might come down to methodological differences between groups. Their conclusions in this paper and in other related papers is that yes, methodology of character delineation really does have an important effect on this problem, particularly with a phylogenetic problem as complex and weakly supported as amphibian origins.
The trees that M&L report are not orthodox. Orthodoxy right now is that lissamphibians originated within amphibamid temnospondyls in the early Permian. M&L do not find that in most cases. The lepospondyl hypothesis has its own history, so it’s not like M&L are suggesting something completely new, but it is not widely accepted. Saying that they are bowing to tradition misrepresents the work that they are doing and, frankly, shows how ignorant you are of what the current state of the field is.
Moreover, M&L are careful to say that the trees they recover are probably not correct in a number of ways, and point out specific things that need to be addressed before we have an early tetrapod phylogeny we can use for broader studies of trends in early tetrapod evolution. Some of those things have been done already and are works in progress or in review. I am optimistic that we’ll have something resembling a stable early tetrapod phylogeny in 4-5 years, and I know some specifics about what that tree will look like. It will likely not resemble “orthodox” phylogenies, but it will also not resemble M&L’s trees in some important ways, nor will it resemble yours.
Thank you.
“Methodology of character delineation” isn’t the only thing we test, though. That’s only one part of testing what happens if everything we’re unhappy with is corrected. Most of these scores are typos and similar mistakes, and most of the rest are scores that are simply outdated because of new publications that came out after 2007 (or 2001…). We find, and report, thousands of such scores. (And I’ve since found a few more.) That’s why Appendix 1 is so extremely long.
It’s really easy to forget which state is called 0 and which is called 1… as we say in the preprint, I’m speaking from experience. I’ve made all the kinds of mistakes myself, except for scoring taxa blockwise.
Thank you for your insights. In summary you appear to be saying the field is still in flux with solutions on the horizon and that I am unaware of the big debates in tetrapod evolution. Okay. I offer results from the data at hand, cognizant that the data can be inaccurate, that convergent traits and crushed specimens make life difficult for systematists.
We’re also saying you’re unaware of issues related to characters: redundant characters (which describe the same things) inevitably distort the resulting tree or at least inflate its support values; sampling too few (independent!) characters is as dangerous as sampling too few taxa; and finally, what you call “the data at hand” are often your misinterpretations of publications, which is a separate question from whether the publications themselves are accurate in the first place.