Earlier the tripod fish, Bathypterois (Fig 1) entered the large reptile tree (LRT, 2228 taxa), but the lack of ossification of so many skull parts made nesting difficult, tentative and ultimately wrong.
Today’s taxon, Aulopus, (Fig 1) was very helpful in nesting the very similar Bathypterois. Adding taxa resolves all phylogenetic problems.
Aulopus filamentosus (Bloch 1792) is the extant royal flagfin, a founding member of the Aulopidae and Aulopiformes, a clade said to resemble lizardfish. Here this bottom dweller (deeper than 1000m) nests with free-swimming Elops, the ladyfish, and Bathypterois, a deep-sea relative.
Figure 1. Aulopus (above) and Bathypterois (below) are close relatives in the LRT.
Bathypterois grallator (Goode and Bean 1886; Sulak 1975; up to 43cm long excluding fin tips) is the extant tripod fish, so named because it stands on extended pelvic and caudal fin tips in the seafloor ooze, facing the current, waiting for tiny planktonic crustacean prey and conspecific mates to come by. If no mates come by, each individual is capable of producing both sperm and eggs. It is derived from Aulopus in the LRT.
This from ScienceDirect.com “In warm surface waters, the sea is practically a saturated solution of calcium carbonate and relatively little energy is needed to maintain calcareous materials (e.g. bone, shells) in solid form. However, solubility rises with increasing pressure and decreasing temperature. In consequence, building and sustaining solid calcareous materials becomes more expensive, particularly at depths beyond 3000 m (below which calcareous sediments are unknown).”
The missing (poorly ossified) skull bones made Bathypterois score in the LRT like an incomplete fossil.
References Bloch ME 1792. Naturgeschichte der ausländischen Fische. Berlin. v. 6. i-xii + 1-126, Pls. 289-323. Goode GB and Bean TH 1883. Reports on the results of dredging under the supervision of Alexander Agassiz, on the east coast of the United States, during the summer of 1880, by the U. S. coast survey steamer “Blake,” Commander J. R. Bartlett, U. S. N., commanding. Bulletin of the Museum of Comparative Zoology.10(5): 183-226. Goode GB and Bean TH 1886. Reports on the result of dredging 38. Description of 13 species and 2 genera of fishes from the “Blake” collection. Bulletin Museum of Comparative Zoology, Harvard 12(5):153–170.
Boesseneckher, Beatty and Geisler 2023 explore every aspect of their new Coronodon (Fig 1) specimens. 27,000+ characters were employed in phylogenetic analysis.
Figure 1. Coronodon, was originally considered a toothed mysticete, but only in the absence of desmostylians, the real ancestors of mysticetes.
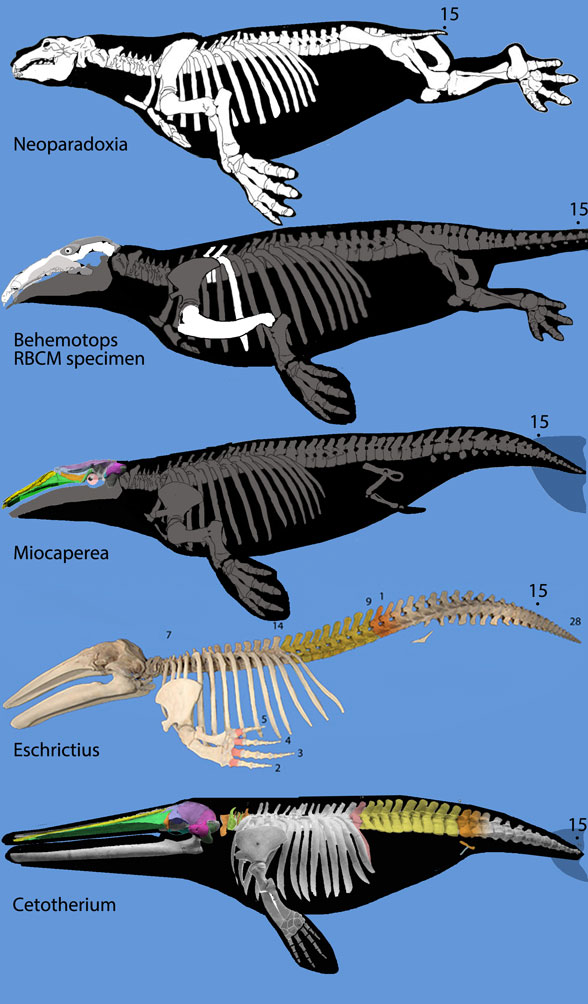
Always good to see a new Coronodon. Unfortunately this study was undercut by taxon exclusion. Adding pertinent taxa nests odontocetes and their archaeocete, pakicetid tenrec and anagalid ancestors far apart from mysticetes and their desmostylian, hippo, mesonychid and oreodont ancestors (Fig 2).
We’ve known this since 2016.
Figure 2. Rorqual evolution from desmostylians, Neoparadoxia, the RBCM specimen of Behemotops, Miocaperea, Eschrichtius and Cetotherium, not to scale.
The authors wrote, “Oligocene fossils have revealed a wealth of early diverging tooth-bearing mysticetes highlighting the transition from archaeocete ancestors to early toothless baleen-bearing eomysticetid whales.”
This is a myth revealed by simply adding taxa. More characters won’t help this paper.
References Boesseneckher RW, Beatty BL and Geisler JH 2023. New specimens and species of the Oligocene toothed baleen whale Coronodon from South Carolina and the origin of Neoceti. PeerJ 11:e14795 DOI 10.7717/peerj.14795
Rietbergen TB et al 2023 described two more of the 30 bats found in the Green River Formation (early Eocene) of Wyoming. They wrote, “The relative stratigraphic position of these fossils indicates that they are the oldest bat skeletons recovered to date anywhere in the world. Taken in sum, our results indicate that Green River bats represent a separate chiropteran radiation of basal bats, and provide additional support for the hypothesis of a rapid radiation of bats on multiple continents during the early Eocene.”
Excellent description. Unfortunately the authors’ outgroups, Sorex (shrew) and Erinaceus (hedgehog), were cherry-picked, are not arboreal and share no bat traits. Also missing is Tadarida, the Mexican free-tailed bat (finger claws lost convergently), which is more primitive than Icaronycteris and Onychonycteris in the LRT.
Just add taxa to solve this problem. Bat sisters are primates and Microcebus is a bat ancestor in the LRT, which minimizes taxon exclusion by adding taxa.
Figure 1. The fruit bat, Pteropus, compared to the small extant bat ancestor, Microcebus.
Several months ago, I suggested (via email) to co-author and bat expert, Nancy Simmons,to add Microcebus (Figs 1, 2). That didn’t work this time. Not sure why bat workers, like pterosaur workers, are reticent to add pertinent taxa to their outgroup lists. There was no reply from Dr Simmons.
Figure 1. An undescribed Green River bat to scale with the skull of Microcebus, the dwarf lemur, an ancestor to bats in the LRT.
Bats likely appeared much earlier than the early Eocene. The early appearance of bats and primates in the placental clade of the large reptile tree (LRT, 2227 taxa) indicates a MUCH earlier evolution of bats, likely in the Jurassic when arboreal placentals, like multituberculates, had free reign to evolve prior to the origin of birds and convergent predatory arboreal theropods, like Microraptor.
References Rietbergen TB, van den Hoek Ostende LW, Aase A, Jones MF, Medeiros ED, Simmons NB 2023. The oldest known bat skeletons and their implications for Eocene chiropteran diversification. PLoS ONE 18(4): e0283505. https://doi.org/10.1371/journal.pone.0283505
It’s a shame when an otherwise fantastic paper hobbles itself with a poor phylogeny.
Werth and Beatty 2023 report, “The presumed “missing link” ancestor to all Cetacea was Indohyus, a small, amphibious digitigrade artiodactyl of the extinct Raoellidae.”
Don’t believe everything you read. Test hypotheses. When you add more taxa to analysis Artiodactyla have nothing to do with mysticete and odontocete origins. Indohyus (Fig 1) was a tenrec sister. Google: Triple Origin of Whales or go to ReptileEvolution.com for details. Better yet, build your own LRT.
Figure 1. Odontoceti (toothed whale) origin and evolution. Here Anagale, Hemicentetes, Tenrec, Indohyus and Leptictidium precede Pakicetus. Maiacetus and Orcinus are aquatic odontocetes.
One other thing… odontocete evolution is characterized by a slow reversal in tooth morphology: 1. complex molars, 2. simple linear crown molars (archaeocetes), 3. all teeth simple cones as in their deep ancestors, the pelycosaurs. These key insights are missing from this paper.
References Werth AJ and Beatty BL 2023. Osteological correlates of evolutionary transitions in cetacean feeding and related oropharyngeal functions. Frontiers in Ecology and Evolution. 11:1179804. doi: 10.3389/fevo.2023.1179804
The extant bowfin (Amia) nests as a semi-basal ray-fin fish in the large reptile tree (LRT, 2227 taxa), a descendant of Middle Triassic Fukangichthys (Fig 1) and Early Triassic Beishanichthys (Fig 1). That answers the headline question.
By contrast, Xu and Gao 2011 described Beishanichthys as a ‘scanilepiform’. “Contrary to previous thought that scanilepiforms are closely related to the Amiidae, the phylogenetic results of this study recognize the Scanilepiformes as stem-group neopterygians. Relationships of the Scanilepiformes and Australosomus with other neopterygians remain unresolved.”
According to Xu and Gao, “The Scanilepiformes Sytchevskaya, 1999 are an extinct group of ‘palaeoniscoid’ fishes with a geological range confined to the Triassic.”
According to the LRT, Beishanichthys is not a palaeoniscoid (= close relatives of Palaeoniscum). Taxon exclusion due to academic tradition is the problem here. The LRT minimizes taxon exclusion by including so many more taxa.
Scanilepis (Fig 3), a recent addition to the LRT prompted by these taxa, nests with Beishanichthys (Fig 1), close to Amia (Fig 1), not related to Palaeoniscum.
Figure 1. Bowfin (Amia) ancestors in the LRT to scale. Here the resemblance between Beishanichthys and Amia is not just superficlal, it is due to a close interrelationship.
Meanhwhile, in their description of Fukangichthys, (Fig 1) Giles et al 2017 reported, “We show that scanilepiforms, a widely distributed Triassic (ca. 251-200 Mya) radiation, are stem polypterids. Polypterids (bichirs and ropefish) represent the earliest-diverging lineage of living actinopterygians.”
By contrast, in the LRT Polypterus is a basal lungfish, not related to Fukangichthys or early diverging actinopterygians (= ray-fin fish). Polypterus is a lobe-fin. Moreover, Scanilepiformes (1999) appears to be a junior synonym for Amiformes (1929).
Unfortunately Prior workers were working under the academic tradition that Amia was a member of the invalid clade, Chondrostei, which traditionally includes bichirs, sturgeons and spoonbills. In the LRT none of these taxa are related to one another. In the LRT Amia, Beishanichthys and Fukangichthys (Fig 1) are closely related basal ray-fin fish (Fig 2).
Figure 2. Extant basal ray-fin fish in the LRT include Engraulis, Malacosteus and Dactylopterus. All three share the trait of a large lateral orbit close to the short rostral tip.
Amia calva (Linneaus 1766; up to 70cm in length) is the extant bowfin, a basal ray-fin fish able to breathe both water and air. As in related Beishanichthys, a single elongate undulating fin is present. Hatchlings look like tadpoles. Fossil relatives of Amia have a worldwide distribution in fresh and salt waters.
Figure 3. Scanilepis from Lehman 1979. Note the differences between the Lehman diagram and the fossil. Colors added here.
Scanilepis dubius (Lehman 1979, Late Traissic, 1.6m long) is a much larger relative of Beishanichthy and the living bowfin, Amia. Note the long dorsal fin and short rostrum. These related taxa indicate the large postorbitals of Amia are the result of fusion between the postorbitals (amber) and jugals (cyan).
This appears to be a novel hypothesis of interrelationships. If not please provide a citation so I can promote it here.
References Giles S, Xu G-H, Near TJ and Friedman M 2017. Early members of ‘living fossil’ lineage imply later origin of modern ray-finned fishes. Nature. 549 (7671): 265–268. Lehman JP 1979. Le genre Scanilepis Aldinger du Rhétien de la Scanie. Bulletin of the Geological Institutio n of the University of Uppsala, N.S 8:113-125. Linneaus C von 1766. Systema naturæ per regna tria naturæ, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Tomus I. Editio duodecima, reformata. pp. 1–532. Holmiæ. (Salvius) . Su T 1978. Memoirs Inst. Vert. Paleont. Paleoanthrop. Peking No. 13. Xu G-H and Gao K-Q 2011. A new scanilepiform from the Lower Triassic of northern Gansu Province, China, and phylogenetic relationships of non-teleostean Actinopterygii PDF. Zoological Journal of the Linnean Society. 161 (3): 595–612. Xu G-H, Gao K-Q and Finarell JA 2014. A revision of the Middle Triassic scanilepiform fish Fukangichthys longidorsalis from Xinjiang, China, with comments on the phylogeny of the Actinopteri. Journal of Vertebrate Paleontology 34(4):747–759.
Pancirolli et al 2016 reported, “The Middle Jurassic docodont Borealestes serendipitus was the first Mesozoic mammal found in Scotland over 40 years ago. Its affinities and morphology have remained poorly understood. We present an updated description and diagnosis of the genus Borealestes, based on high-resolution micro-computed tomography (micro-CT) and synchrotron scans. Our phylogenetic analysis supports a clade formed by Borealestes, Haldanodon, Docofossor, and Docodon. Ontogenetic variation in the mandibular morphology of Borealestes is similar to that seen in Docodon and Haldanodon, with the delayed emergence of the ultimate lower molar, the shift of the last molar to the front of the coronoid process, and a posterior shift of the Meckel’s sulcus in successively older individuals. This supports a distinctive growth pattern in the clade including Borealestes and Docodon, one that may be present in Docodonta as a whole.”
Borealestes and Haldanodon are not mammals according to the large reptile tree (LRT, 2226 taxa). The were late-surviving [Jurassic] probainognathid cynodonts that independently developed mammal-like teeth (Fig 1) and lots of them in tiny Borealestes.
Figure 1. Borealestes restored and mandible from Panciroli et al 2016. Tooth colors added here. The authors indicated a double-rooted molar, here re-identified as a two-rooted worn down first premolar.
According to Wikipedia, “Docodonta is an order of extinct mammaliaforms that lived during the Mesozoic, from the Middle Jurassic to Early Cretaceous. They are distinguished from other early mammaliaforms by their relatively complex molar teeth.”
The putative clade, ‘Mammaliaformes‘ is a junior synonym for ‘Mammalia’ since it relies on the basal marsupial, Morganucodon, as an outgroup.
Middle Triassic Prozostrodon (Fig 2) was not tested, nor mentioned in the text.
The most interesting thing about this taxon is the parallel evolution of mammal-like teeth into the Middle Jurassic. There were more teeth than any mammal. Here the double-rooted canine is reinterpreted as the first of six premolars. The single-rooted canine was probably no larger than the incisors, based on its diameter.
Figure 2. The cappa-ufsm specimen of early Late Triassic Prozostrodon compared to size and to scale with two specimens of the smaller Middle Jurassic Borealestes.
Panciroli defines mammals with lactation and three inner ear bones, rather than the preferred last common ancestor method (e.g. Megazostrodon, Ukhaatherium, their last common ancestor and all of its descendants).
Panciroli gives herself credit for moving that origin of mammals to 250mya (in the video) to 300 mya (in the caption). This is not news.
According to the author, the first group to split off from ‘the amorphous taxa’ were the amphibians (= the anamniotes according to the caption). She says, “Amniotes laid eggs with a tough, leathery shell.” Not necessarily. The shell is another matter. The amnion is the key membrane.
More textbook mythology follows. The author reports the first split was between the synapsids and diapsids. This is wrong on at least two levels. All basal reptiles had no skull openings. Diapsid architecture evolved twice, once from synapsid ancestors.
The author thinks the story of pelycosaurs “is missed out of our telling, not just of mammal history, but really history of life on Earth.” She focuses on Dimetrodon.
This is wrong. Dimetrodon is not part of the mammal story. We knew this thirty years ago. She claims Dimetrodon is not a reptile. This is wrong. All animals with an amniotic membrane are reptiles. Where is the concept of monophyly? Dr. Panciroli is stuck in the 1960s.
Next Panciroli describes therapsids as the most diverse lineage in the Permian, wasting time with dicynodonts. This is supposed to be the untold story of mammal evolution. She mentions scale-less skin preservation in Estemmenosuchus, not realizing that all early amniotes and many Permian taxa lacked scales plesiomorphically. Scales appeared in the Triassic, spurred by the rise of biting insects.
After 34 minutes Panciroli discussed her first mammal ancestor, Procynosuchus.
At 35 minutes Dr. Panciroli describes Thrinaxodon, with a complex and branched maxillary canal, indicative of whiskers. Again, this information is at least thirty years old. Panciroli attributes milk production to Thrinaxodon, because it loses the pineal opening. This is wrong. Single replacement teeth indicate milk production. So does the last common ancestor method, which attributes milk production to nothing before monotremes.
Panciroli reported mammals begin to evolve smaller sizes during the Mesozoic. This is only occasionally true (e.g. Hadrocodium), but most early mammals tested in the LRT were mouse-to squirrel-sized. I only hope the book is better than the teaser talk.
Correctly Panciroli describes the reduction of posterior jaw bones to ear bones. This is not news.
At 41:00 Panciroli acknowledges in general terms a diverse assemblage of Mesozoic mammals, none larger than a badger this time.
At 43:00 the author describes Early Cretaceous Repenomamus, not realizing it was a late-survivng, pre-mammal cynodont, not a mammal.
At 45:00 Panciroli discusses the asteroid at the K-Pg boundary. So out of 50 minutes, only ten were given to the ‘untold’ Mesozoic mammal story. No time was given to the Cenozoic taxa. For the remaining 32 minutes viewer questions were answered.
Suggestion to Dr. Panciroli: Build your own LRT so you can understand the evolution of mammals and their untold story. It’s not good to cherry-pick a few well-worn tales published years ago and tell readers, your book is the ‘untold story’.
References Pancirolli E, Benson R and Luo Z-X 2016. The mandible and dentition of Borealestes serendipitus (Docodonta) from the Middle Jurassic of Skye, Scotland. Journal of Vertebrate Paleontology 39(3):Article: e1621884 Waldman M and Savage RJG 1972. The first Jurassic mammal from Scotland”. Journal of the Geological Society. 128 (2): 119–125.
The longest lived minor clade of placental mammals, Multituberculata (Middle Jurassic to Late Eocene), mysteriously went extinct when similar placentals did not. Perhaps this was due to the rise of carnivorous raptorial birds (e.g. hawks, eagles, owls).
This is a novel hypothesis based on timing and hearing.
Figure 1. Animation of the mandible of the multituberculate Kryptobaatar showing the sliding of the jaw joint producing separate biting and grinding actions, just like rodents, their closest relatives in the LRT.
As long-time readers might recall multituberculates evolved from rodents and plesiadapiformes in the large reptile tree (LRT, 2225 taxa), the only cladogram to test and nest these three clades together.
Due to the sliding jaw joint of multis (Fig 1) their three tiny ear bones failed to develop into the exquisitely sensitive tiny middle ear bones of their relatives. Chalk that up to a reversal via neotony. Their post-dentary bones stopped developing at the embryo stage and remained large elements attached to the dentary as an adult, like those of pre-mammals. Many arboreal therians recapitulate phylogeny during ontogeny like this.
That’s the only reason why many paleontologists consider multis to be at least as primitive as monotremes, if not more primitive. That’s why they don’t test multis with rodents and plesiadapiformes. The LRT is not limited by such assumptions.
Figure 2. Comparing multituberculate origins: Cziki-Sava et al. vs. LRT.
Poor hearing was fine so long as multis had the trees largely to themselves. However, by the Eocene a new clade of predators capable of snatching arboreal mammals out of the branches came along.
Figure 3. The aye-aye, Daubentonia in vivo. This is the closest living relative of multituberculates and is itself a plesiadapiform member of Glires, close to rodents, not primates.
Hawks didn’t care whether their prey was a rodent, a plesiadapiform, a primate or a multituberculate. Hawks attacked them all. Unfortunately multis were then at a disadvantage, due to their less acute hearing. As a result they were relatively ‘easy pickins’ and their numbers dwindled to extinction. Only rodents, with their excellent hearing, managed to survive worldwide. Small primates (including lemurs and adapids) disappeared except in heavily jungled areas near the equator. Plesidapiformes survived as a single nocturnal jungle taxon, Daubetonia, the aye-aye, on Madagascar, along with native (not rafting) lemurs. Owls also hunt on Madagascar, so it wasn’t the mere presence of nocturnal hunting birds that doomed arboreal mammals. It was the newly evolved hunting birds > plus < the less acute hearing of multis that hastened their undoing.
Or is there a better explanation? Let me know if you have one.
Van Valen and Sloan 1966 wrote: “The multituberculate mammals declined in numerical abundance from the late Cretaceous to their extinction in the late Eocene. Their maximal diversity, however, occurred in the late part of the middle Paleocene, suggesting an increase in specialization in the face of competition from placental mammals, which diversified very rapidly in the Paleocene. Various lines of evidence lead to the conclusion that first condylarths, then primates, and finally rodents contributed to the gradual extinction of multituberculates, at least in part by evolution in the same region of competing resource requirements.”
Maas, Krause and Strat 2016 echoed Krause 1986 when they wrote: “Analyses of body size, diet, activity patterns, and locomotion support the hypothesis that rodents and non-paromomyid plesiadapoids may have competed for the same resources.”
Like other workers, these authors do not mention poor hearing and hawks.
References Krause, DW 1986. Competitive exclusion and taxonomic displacement in the fossil record: the case of rodents and multituberculates in North America. Pp. 95–117. In Flanagan, K. M., and Lillegraven, J. A. (eds.), Vertebrates, Phylogeny and Philosophy. Contributions to Geology Special Paper 3. The University of Wyoming; Laramie, Wyoming. Maas M, Krause DW and Strait SG 2016. The decline and extinction of Plesiadapiformes (Mammalia: ?Primates) in North America: displacement or replacement? Paleobiology 14(4): Van Valen L and Sloan RE 1966. The extinction of the multituberculates. Systematic Zoology 15:261–278.
Burin et al 2023 looked at mysticete and odontocete body length, then added fossil taxa to document trends in ‘whale’ size.
Figure 1. Odontoceti (toothed whale) origin and evolution. Here Anagale, Hemicentetes, Tenrec Indohyus and Leptictidium precede Pakicetus. Maiacetus and Orcinus are aquatic odontocetes. This is an older illustration. Adding taxa moved Andrewsarchus and Sinonyx away from this clade.
Figure 2. Taxa in the lineage of right whales include Desmostylus, Caperea and Eubalaena. The tiny bit of jugal posterior to the orbit (in cyan) is found in all baleen whales tested so far. The frontals over the eyes are just roofing the eyeballs in Desmostylus, much wider in Caperea and much, much longer in Eubalaena.
According to phys.org/news: “To get a more detailed look at cetacean evolution [ there is no such thing as a ‘cetacean’], a team of scientists, including Dr. Travis Park and Dr. Gustavo Burin, both Leverhulme postdocs at the Natural History Museum, gathered body length measurements of 345 different species, including 89 living species and 256 fossil lineages, in the largest dataset of its kind. Their article, “The Dynamic adaptive landscape of cetacean body size” is published in Current Biology.”
“By comparing body lengths from both living and fossil species—an approach rarely taken—the research revealed that the evolutionary trends in cetacean size remained much the same for over 20 million years after they entered the ocean.”
Mysticetes and odontocetes entered the ocean separately.
“The first cetaceans were goat-sized animals which lived on the edge of lakes and rivers, spending time both in and out of water. Over time, their descendants became increasingly adapted to life in water, before leaving land behind altogether.”
Since these authors have no idea that mysticetes evolved by convergence with mysticetes, this paper becomes a waste time for the writers, editors, referees and readers – unless it is split into two works. Readers, make sure your phylogeny is valid.
References Gustavo Burin et al 2023. The dynamic adaptive landscape of cetacean body size, Current Biology (2023). DOI: 10.1016/j.cub.2023.03.014
The earliest (= most primitive) bony fish had a jugal (cyan in figure 1), but no squamosal (magenta in figure 3) tetrapod homolog. The squamosal first appeared in flat-head, pre-tetrapod fish like Grossius and Tinirau (Fig 3) between splitting jugal elements that were a large single element in Eusthenopteron (Fig 2).
Figure 1. Tarrasius is a small Early Carboniferous fish basal to extant moray eels. Note the presence of a jugal (cyan), but a lack of a squamosal (magenta cheek bone) as seen in figure 2.
The recognition of the splitting of the jugal with the appearance of the squamosal is one of those vexing things that made this phylogenetic study of fish extend as long as it has. This was vexing because traditional labels changed.
Figure 2. Eusthenopteron was originally labeled with a squamosal (cyan). Here that bone is a large jugal and a squamosal is not present. The two tan bones are here labeled lacrimals. The posterior lacrimal is on its way out after original detachment from the maxilla (see figure 1). The tetrapod lacrimal (anterior tan element here) is from the circumorbital ring, originally all postorbital elements (amber). Compare to figure 1. The anterior lacrimal migrates closer to the posterior naris, a pre-choana that migrates to the jaw margin, then moves on to the palate, disappearing from the exterior in pre-tetrapods. Compare this skull to Selenodus in figure 3. The lacrimal precedes the premaxilla and maxilla, forming a base for both to develop on in certain placoderms, sharks and acanthodians.
Ray-fin fish never developed a squamosal and neither did many lobe fins. Starting with Late Devonian Grossius, a lobe-fin with a wider-than-tall skull (Figs 3, 4), a squamosal first appears and is largely retained in tetrapods.
Figure 3. Left: Grossius demonstrates the origin of the squamosal (magenta), splitting the twin jugals. The upper jugal eventually disappears. Right: Selenodus shows the plesiomorphic (pre-squamosal) condition. Schultze 1973 thought both of these taxa were related to Onychodus. In the LRT only Selenodus is related to Onychodus.
Grossius aragonensis (Schultze 1973; Late Devonian) has a straight rostrum and a wider-than-tall skull. Note the splitting of the jugal as the squamosal (magenta) moves the two jugal elements apart.
Figure 3. Grossius compared to Middle Devonian Tinirau, a flatter pre-tetrapod, pre-Panderichths taxon. The splitting of the jugal is just one of many things you learn > after < studying fish skulls for 5 months. Sorry it took so long. I started as a freshman on this topic and the labels were misleading.
Tinirau clacke (Swartz B 2012; Middle Devonian; UCMP 118605) nests between Cabonnicthys and Panderichthys in the LRT. The skull is twice as wide as tall, the skull bones are more tetrapod-like, dorsal ribs make a first tentative appearance. The tail was likely straight, as in sister taxa.
This hypothesis re-labels fish skull bones with tetrapod homologs to facilitate tracing their development across all clades and taxa (Figs 1–3) in the LRT. I wish this practice was more widespread. Traditional academics also label many fish skull bones this way, but not to this extent. They have their own nomenclature.
For example: the traditional fish preorbital is a tetrapod homolog postorbital (the entire circumorbital ring, other than the postfrontal). In one clade a portion becomes the tetrapod lacrimal. Other examples are seen in figure 2.
The preopercular and opercular are not found in tetrapods, nor are they transformed during evolution into new bones. They just disappear.
This appears to be a novel hypothesis of fish skull bone homologs. If not, please provide a citation so I can promote it here.
References Schultze H-P 1973. Crossopterygier mit heterozerker Schwanzflosse aus dem Oberdevon Kanadas, nebst einer Beschreibung von Onychodontida-Resten aus dem Mitteldevon Spaniens und aus dem Karbon der USA. Palaeontographica, Abteilung A, 143:188–208. Swartz B 2012. A marine stem-tetrapod from the Devonian of Western North America. PLoS ONE. 7 (3): e33683. doi:10.1371/journal.pone.0033683
Amaral et al 2013 described Sapperichthys (Fig 1) as a new gonorynchid from the Cenomian (earliest Late Cretaceous) of Chiapas, Mexico. The authors did not consider sturgeons in their phylogenetic analysis, nor did they mention sturgeons in their text.
Figure 1. Sapperichthys from Amaral et al 2013. Rostrum and tail lost during excavation. Colors and reconstruction added here. In situ fossil shown about actual size on 72dpi monitors. Note the diamond-shaped dorsal armor in the fossil, as in Acipenser. See figure 2.
Figure 2. Acipenser brevirostrum, 1m typical length. Records up to 1.47m.
This appears to be a novel hypothesis of interrelationships overlooked due to taxon exclusion. If not please provide a citation so I can promote it here.
References Amaral CRL, Alvarado-Ortega J and Briot PM 2013.Sapperichthys gen. nov., a new gonorynchid from the Cenomanian of Chiapas, Mexico. In: Mesozoic Fishes 5 – Global Diversity and Evolution, Arratia, Schultze and Wilson (eds.): 305–323. by Verlag Dr. Friedrich Pfeil, München, Germany – ISBN 978-3-89937-159-8