Until yesterday it was traditionally thought and taught that labial cartilages (hinged elements lateral to the jaws of some sharks and kin, Figs 1–6) appeared de novo (= without precedent or homolog).
Figure 1. Mako shark (Isurus) with a focus on the labial cartilage and quadrate (red).
Klimpfinger and Kriwet 2020 reported, “We identified labial cartilages in representatives of 27 from 36 shark families and detected different numbers, sizes and forms of labial cartilages, as well as differing positioning along the jaws.”
Figure 2. Daliatias, the extant kitefin shark, has relatively large labial cartilages (red). The jugal homolog (cyan), usually fused, is not fused here.
Now it looks like labial cartilages are homologs of (and therefore derived from) the external quadrate plate on placoderms like Coccosteus (Fig 3). Sometimes labial cartilages shrink and disappear. Most bony fish and tetrapods lack labial cartilages. In other taxa labial cartilages can enlarge and migrate (Fig 2).
Figure 3. Xiushanosteus and Coccosteus demonstrates the origin of the quadrate (red), which splits to form an external labial cartilage and an internal typical quadrate.
Gregorius rexi is a basal chondrichthyan that has a primitive labial cartilage (Fig 4 red angled arc). Unfortunately, due to taphonomic scattering, the exact location of this element in this specimen is difficult to determine. So in the reconstruction (Fig 4) the external quadrate = labial cartilage is located close to the internal quadrate based on comparative anatomy to Coccosteus (Fig 4).
Figure 4. Gregorius rexi is a Carboniferous chondrichthyan with a Silurian origin. Here the external quadrate-labial cartilage is small, not articulated and shown in red. Compare to Coccosteus in figure 3.
Turns out that other placoderm descendants, like catfish (Figs 5, 6), also retain an internal and external quadrate (= labial cartilage), in some, but not all from this clade.
Figure 5. The catfish Diplomystes with a focus on the internal quadrate and external quadrate (= labial cartilage).
Here the catfish labial cartilage homolog in Dipolomystes(Fig 5) is shown in red.
This appears to be a novel hypothesis of homology. If not, please provide a citation so I can promote it here.
References Klimpfinger C and Kriwet J 2020. Comparative morphology of labial cartilages in sharks (Chondrichthyes, Elasmobranchii). The European Zoological Journal 87(1):741–753.
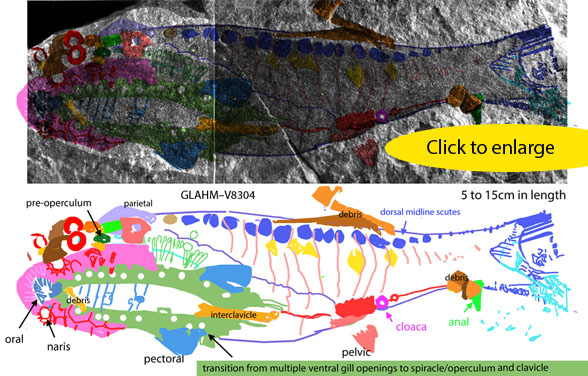
Early Devonian Doliodus (Fig 1) and Early CretaceousCalamopleurus(Fig 1) are finally united in the large reptile tree (LRT, 2308 taxa). Re-identifying the postorbital bones brought these two long-tooth taxa together. Note the dorsal views.
Doliodus was long considered a chondrichthyan, but has branchials located between its mandibles, like a bony fish.
Figure 1. Early Devonian Doliodus and Early Cretaceous Calamopleurus, not to scale. These two nest together in the LRT.
Doliodus problematicus (Whiteaves 1881; Traquair 1893; Miller, Cloutier and Turner 2003; Maisey et al. 2018; Early Devonian) was considered the ‘oldest articulated chondrichthyan’ and a transitional taxon between acanthodians and sharks. Here Doliodus nests with Calamopleurus and Sauropsis. The skull is wider, the maxilla is flat and the dual-pronged teeth are oriented medially). This was probably a nocturnal bottom feeder. Like spiny sharks the leading edge of the pectoral fin was a spine.
This appears to be a novel hypothesis of interrelationships. If not, please provide a citation so I can promote it here.
References Maisey JG et al. (6 co-authors) 2018.Doliodus and Pucapampellids: Contrasting perspectives on stem chondrichthyan morphology. Chapter 5 in Evolution and Development of Fishes. Miller RF, Cloutier R and Turner S 2003. The oldest articulated chondrichthyan from the Early Devonian period. Nature 435:501–504. Traquair RH 1893. Notes on the Devonian fishes of Campbellton and Scaumenac Bay in Canada. No. 2. Geological Magazine, decade 3, 10: 145–149. Turner S and Miller RF 2004. New ideas about old sharks. American Scientist 93:244–252. Whiteaves JF. 1881. On some fossil fishes, Crustacea and Mollusca from the Devonian rocks at Campbellton, NB, with descriptions of five new species. Can Nat 10:93–101.
The flat Late Devonian placoderm with tiny lateral eyes, Cowralepis (Fig 1), is known from a growth series of several individual specimens. Here both its dorsal and ventral views are exposed. Yes, these specimens look wrinkled and deflated. What does that mean for a placoderm? I wonder if their bones were not as well ossified? They look like shed skin, but that’s not how placoderms matured.
Moving on…
Figure 1. Cowralepis in dorsal and ventral views. Here the dislocated pemaxillae are returned to their in vivo positions.
After re-analysis in DGS using an updated Drepanolepis Bauplan (Fig 2) Cowralepis (Fig 1) is now better understood. Note the displaced premaxillae here returned to their in vivo positions. Those tooth plates were overlooked earlier. Their presence changes scores.
Figure 2. Drepanolepis. Colors added here. Compare to Cowralepis in figure 1. Note the post-crania here is not represented by a thoracic carapace and plastron, but by heavy armor.
Cowralepis mclachlani (Ritchie 2005; Carr, Joahnson and Ritchie 2009. Middle Devonian) is a phyllolepid placoderm derived from Entelognathus. The skull is disc-like with a ventrally open premaxilla and transverse mandible with a lateral articular and quadrate, hearkening back to Qilinyu.
Is this a reversal?
References Carr RK, Johanson Z and Ritchie A 2009. The phyllolepid placoderm Cowralepis mclachlani: insights into the evolution of feeding mechanisms in jawed vertebrates. J Morphol 270(7):775-804. doi: 10.1002/jmor.10719. Ritche A 2005. Cowralepis, a new genus of phyllolepid fish (Pisces, Placodermi) from the Middle Devonian of New South Wales. Proceedings of the Linnean Society of New South Wales 126:215–259.
Yesterday we looked at four placoderm taxa (Fig 2) that seemed to be related and seemed to lose a gracile, bowed mandible in an evolutionary process based on the data, but imagined here based on comparative anatomy.
Good news… the four taxa nested together in the LRT with yesterday’s data. That initial guess was tested and confirmed, now awaiting other workers repeating this experiment.
Figure 1. Dickosteus in situ. DGS colors and reconstruction added here. Note the prognathus mandible anchors on the red quadrate. Note sure what the pink-green-cyan strip represents, possible male organs of intromission.
Today a DGS tracing of Dickosteus (Fig 1) revealed a premaxilla and prognathus mandible not presented in the original graphic (Fig 2), which also omitted the thoracic shield and caudal bones.
This affirms the earlier guess (Fig 2) that related Homosteus and Yinostius likely also had a prognathus mandible despite a lack of evidence for any mandible in the available data.
Figure 2. A series of placoderm taxa that appear to lose a gracile mandible based on the data. Dickosteus in figure 1 provides evidence that a prognathus mandible was present at least in that taxon.
Dickosteus threiplandi (Miles and Westoll 1963; Middel Devonian) is similar to Coccosteus with a distinct suture pattern. Here (Fig 1) a specimen is traced in DGS colors and reconstructed revealing a premaxilla and mandible not presented in the original graphic. The mandible attaches to the quadrates (red), pushing it out ahead of the rostrum. During taphonomy the mandible was backshifted below the skull, perhaps because the jaw joint was weak in this clade.
References Miles RS and Westoll TS 1963. Two new genera of coccosteid Arthrodira from the Middle Old Red Sandstone of Scotland, and their stratigraphical distribution. Trans. R. Soc. Edinb, 65, 179–210.
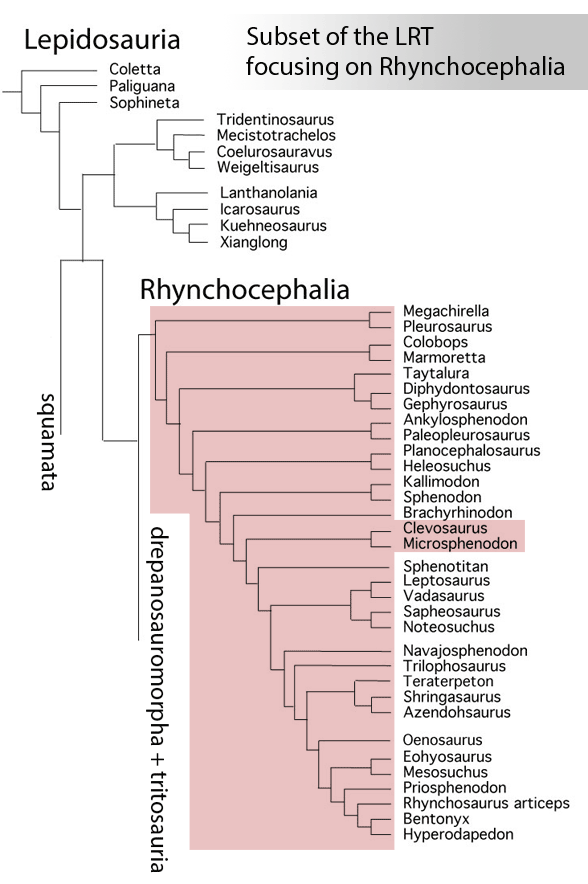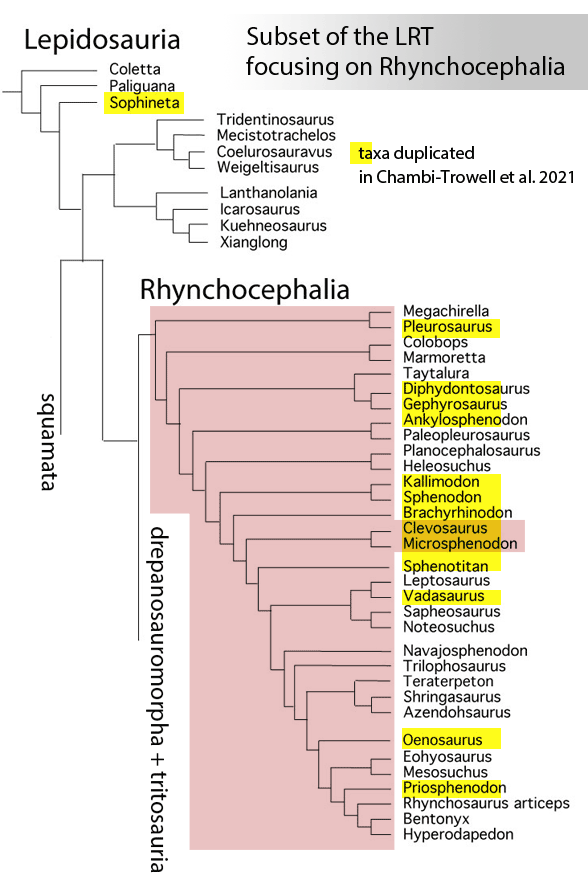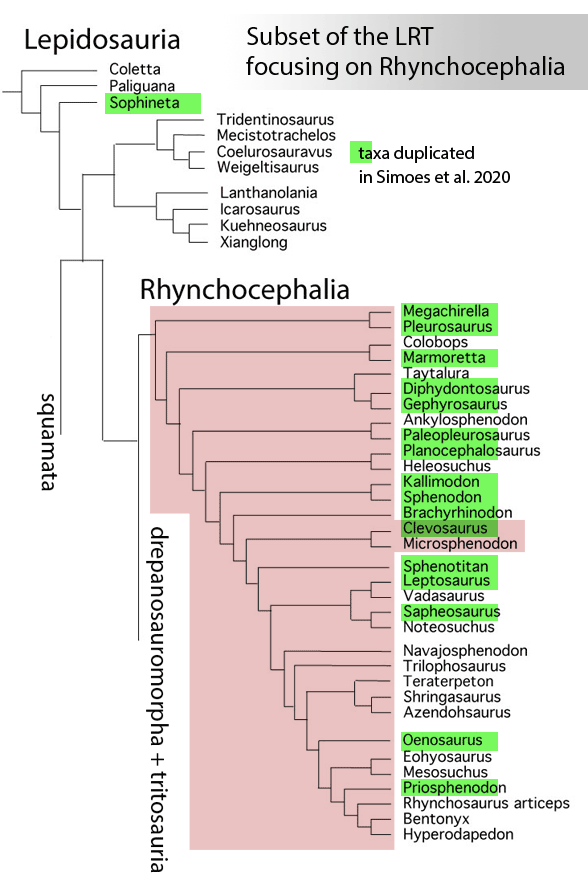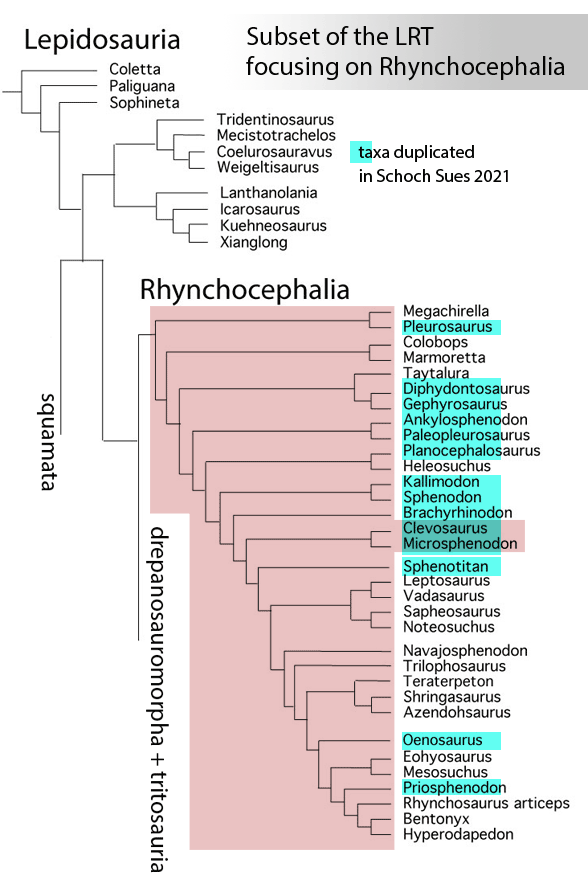
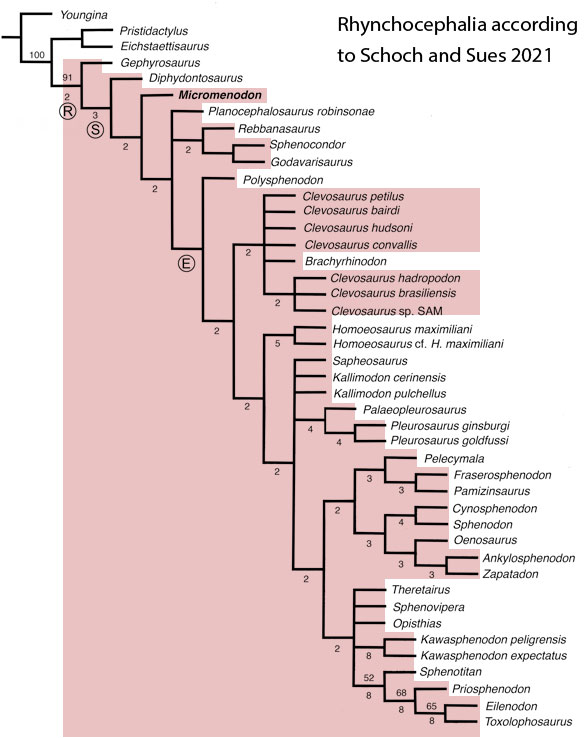
Sues and Schoch reported on “Wirtembergia hauboldae, the oldest rhynchocephalian known to date… Phylogenetic analysis recovered the new rhynchocephalian as the earliest-diverging member of its clade.”
The authors datedWirtembergia to the Ladinian, Middle Triassic. Wirtembergia is also a province in southern Germany (aka Baden-Württemberg). This paper was behind a paywall at this time. Whenever your taxon nests outside the clade you’re trying to nest it in, consider adding taxa.
Figure 1. Wirtembergia compared to Sphenodon, Kudnu and Carbonodraco. Note the large lateral naris not seen in the rhynchocephalian, Sphenodon.
From the Sues and Schochabstract: “Premaxilla with four teeth, first being largest and decreasing in size from first to fourth. Jugal with tiny, spur-like posterior process. Lateral surface of dentary strongly convex dorsoventrally for much of length of bone, bearing distinct longitudinal ridge and sculpturing in large specimens. Coronoid eminence of dentary low, subrectangular, and with dorsoventrally concave lateral surface in larger specimens. Dentition with pleurodont anterior and acrodont posterior teeth. Posterior (=additional) teeth with (in side view) triangular, at mid-crown level labiolingually somewhat flattened crowns, and with oval bases.”
As reported, the specimen (Fig 1) consists of a premaxilla, jugal and dentary
After analysis, Schoch and Sues nested Wirtembergia as the outgroup to Gephyrosaurus + Rhynchocephalia. Several protosquamates (e.g. Sophineta, Marmoretta) and two archosauromorphs (Youngina, Prolacerta) were cherry-picked as outgroup taxa.
When Wirtembergia was added to the large reptile tree (LRT, 2308 taxa, subset Fig 2) it nested outside the Rhynchocephalia. Wirtembergia shared more traits with Kudnu and Carbondraco (Fig 1), two early lepidosauromorphs with a large lateral naris, a tall maxilla, large anterior premaxillary teeth and a down-tipped dentary, among a longer list of similar traits. These three nest basal to Stephanospondylus + pareiasaurs + hard shell turtles + soft shell turtles. Outgroup taxa include diadectids, which helps explain the large blunt anterior teeth.
Figure 2. Subset of the LRT focusing on the Rhynchocephalia. Here the earliest member is Mesosuchus, a derived taxon basal to Late Triassic rhynchosaurs.
PS In the LRT the earliest known member of the Rhynchocephalia is Early Triassic Mesosuchus (Fig 2) a taxon more derived than any sphenodontian. Schoch and Sues had no trouble adding Priosphenodon to their 2021 cladogram. but omitted Mesosuchus, which they considered an unrelated basal rhynchosaur. Add taxa to find out for yourself.
References Sues H-D and Schoch RR 2023. The oldest known rhynchocephalian reptile from the Middle Triassic (Ladinian) of Germany and its phylogenetic position among Lepidosauromorpha. The Anatomical Record. https://doi.org/10.1002/ar.25339
Payne, Mannion, Lloyd and Davis 2023 reported, “abiotic and biotic drivers shaped 250 million years of diversity in crocodile-line archosaurs.”
“The phylogeny was generated via a ‘metatree’ approach93. This approach is similar to formal supertree analysis but differs in that the input is not published trees but the original character matrices or sequence alignments that are themselves reanalysed to generate more complete sets of source topologies.“
That means the authors did not have to look at specimens or photos of specimens… but taxa were not the focus here…
“We also included a molecular tree containing 23 extant taxa. This tree was upweighted in the final matrix to account for the disproportionate influence of morphology on the position of Gavialis gangeticus. The analysis found 1,320 MPTs.”
That’s a lot of MPTs. Not sure about using molecules for extant crocs, which likely had a deep time divergence.
On the publicity front, from Phys.org “Researchers map crocodile family tree to shed light on their evolution”
“A research team, led by scientists at the University of York, mapped the family tree of the ferocious ambush-predators and their extinct relatives known as Pseudosuchia. They then compared this with data from the fossil record to understand why crocodiles have so few living species, while there are 11,000 species of their closest living relatives, birds.
A competing analysis (the LRT) does not recognize a monophyletic ‘Pseudosuchia‘. Instead, only Crocodylomorpha + Dinosauria = Archosauria in the LRT. Oddly, the basal dinosaur, Herrerasaurus, is not in the taxon list. It’s an ideal outgrourp taxon for crocs. To their credit the authors included lots of poposaurs, the outgroup clade to the Archosauria in the LRT.
The published cladogram (Fig 1) has green and orange lines instead of taxon names and only silhouettes of a few icons to tell the reader about what they are looking at.
Figure 1. Closeup of the cladogram from Payne et al. 2023. Instead of taxonomic names, color bars are used in this paper.
Phys.org continues: “The researchers found that climate change and competition with other species have shaped the diversity of modern-day crocodiles and their extinct relatives, but the findings also reveal ecology—whether species live in the sea, in freshwater or on land—played an unexpectedly key role in survival.”
“The study found that when global temperatures rose, the number of species of the crocodile’s sea-dwelling and land-based relatives went up, while increases in competition for resources, perhaps with sharks, marine reptiles or dinosaurs, likely brought about their extinction. By contrast, the crocodile’s freshwater-dwelling relatives were not affected by temperatures, but were put at greatest risk of extinction by rising sea levels.”
As you can see (Fig 1), individual taxa were not the focus of this study, which was more about numbers, niche, weather and deep time.
Figure 2. Random portion of the data matrix from Payne et al 2023. Gray bar represents my selection of Lotosaurus as a taxon.
Unfortunately, the published data matrix (suppdata) would not open in PAUP 4.0a. Lots of taxa were scored with lots of “??” (Fig 2). Taxa missing from the study include: Scleromochlus, Saltopus, Faxinalipterus, Lewisuchus and Dyoplax.
References Payne ARD, Mannion PD, Lloyd GT and Davis KE 2023. Decoupling speciation and extinction reveals both abiotic and biotic drivers shaped 250 million years of diversity in crocodile-line archosaurs. Nature ecology and evolution. https://doi.org/10.1038/s41559-023-02244-0
Just wondered if more data could be pulled out of one of this remarkable soft-tissue specimen of the placoderm with dentary teeth, Late Devonian Amazichthys (Figs 1, 2). Jobbins et al 2022 published the photo and diagram. Such preservation provides rare insight. The diagram was step one. This is step two.
Step three would be a larger, more detailed treatment performed by a young someone with access to the fossil eager to write a dissertation.
Figure 1. Amazichtys from Jobbins etal 2022. DGS colors added here. This appears to be dorsal exposure, rather than the ventral one in the authors’ diagram. A mold and a cast present the same data only different. There is debris around this fossil, but who’s complaining? This is remarkable preservation and presentation.
Amazichthys trinajsticae (Jobbins et al 2022; Late Devonian, Figs 1. 2) was preserved with a body outline indicating a deep caudal and dorsal fin. Otherwise this placoderm was a larger version of tiny Millerosteus in most respects, including small sharp dentary teeth!
Figure 2. Amazichthys skull in situ. Colors added here.
For those interested in imagining the soft-tissue post-crania of Dunkleosteus, this specimen (Fig 1) is the place to start. Note the size of the skull vs everything else.
References Jobbins M, Rücklin M, Ferrón HG and Klug C 2022. A new selenosteid placoderm from the Late Devonian of the eastern Anti-Atlas (Morocco) with preserved body outline and its ecomorphology. Frontiers in Ecology and Evolution 10:969158. doi: 10.3389/fevo.2022.969158
In order to know taxa better it’s worthwhile to revisit earlier interpretations once a bit of experience working with clade patterns is gained. The is the next step beyond freshman naiveté. In this case, several taxa (Figs 1–4) were revisited after earlier interpretations beset with first-timer mistakes. The resulting new interpretationsupdate and correct prior errors and improve analytical scores.
Figure 1. Tiny roadkill Shenacanthus now has a new skull with a gracile mandible and other changes. Ozarcus has a palatine strip (cyan) rather than the former maxilla misinterpretation. No taxa in the shark line develop a maxilla. The long hyomandibular (dark green) was broken in two during taphonomy. It is reassembled here with the crack line preserved.
Early Silurian Shenacanthus (Fig 1, Zhu et al 2022) has a 5mm skull crushed and somewhat scattered. Here, after a bit of experience and sweat equity, Shenacanthus is re-reconstructed. Now it is even more similar to the much larger, much later big-eyed basal shark, Ozarcus.
Early Carboniferous Ozarcus (Fig 1, Pradel et al 2014) was originally µCT scanned, so it is perfectly presented. Earlier I mistook the strip-like palatine for a maxilla. This simple correction was made by changing its DGS color to cyan. Now no chondriichthyans have a maxilla.
Figure 2. Thelodus has new DGS colors here and these move thelodus closer to sturgeons in the LRT. Several pieces of debris are identified. Click to enlarge.
Silurian Thelodus (Fig 2) is a dark on dark 2D fossil that was difficult to sort out and interpret. Here, after some experience and several stumbling attempts DGS colors now indicate this specimen was basal to sturgeons with small fins and ventral nares. This linking Thelodus to heterostracans, and indicates their progress in their migration to their eventual position anterior to the orbits in extant sturgeons. A tiny hyomandibular disc (dark green) makes an appearance near the spiracle, prior to its enlargement as yet another type of operculum. FInally, if not yet another piece of debris, the interclavicle makes an appearance at the flat ventral midline of Thelodus. This bone is retained in later placoderms, which underlap it with lateral elements.
Figure 3. The skull of Devonian Euphanerops, a relative of Bikenia in the LRT. And a relative of the conodont, Promissum. All three are early gnathostomes in the tetrapod line. Here the maxillae arc (sans premaxilla, which appears later) and the straight dentary elements are pulled out to reveal their identities.
Devonian Euphanerops, (Fig 3) is a lamprey-like, lancelet-like (Fig 4) relative of Bikeniaand the conodont, Promissum in the LRT. Like conodonts, Euphanerops had ‘jaws’ made up of several separate slender elements which linked more tightly together in more derived gnathostomes basal to tetrapods. Here the maxillae form an arc without a premaxilla. By contrast, the early dentary elements were straight. Conodonts can have a dozen of these, but Euphanerops has fewer. Here all the jaw elements lack teeth or conodont-like sharp processes.
Figure 4. Not two unusual anal fins, but two typical pelvic fins appear in Euphanerops as shown here. The atrium was misidentified as gut and anus by Sansom, Gabbott and Purnell 2013.
The pelvic fins of Devonian Euphanerops, (Fig 4, Woodard 1900; Janvier and Arsenault 2007) were identified as unusual paired anal fins by Sansom, Gabbott and Purnell 2013 because the atrium excurrent valve was miisidentified as the anus (= the cloaca). Here corrections are offered in DGS colors. So far I have not seen any paired anal fins.
Figure 5. Traced here using DGS methods reveals overlooked vestigial fins in Carboniferous Paratarrasius.
All the fins on the Carboniferous eel-like Paratarrasius How would you score this taxon? Did it have a straight tail? Or a hetercercal tail? Comparie Paratarrasius (Fig 5) to its LRT sister, Bluefieldius (Fig 6), which has a heterocercal tail. That makes the dorsal fin of Paratarrsius extend to the tail tip (not beyond as the diagram (Fig 5, lower right) shows. In addition, pelvic and anal fins were originally overlooked based on the finless diagram. The pectoral fins were imagined with dashed lines to look like ping-pong paddles. Here all laferal fins are vestigial, but still present. They are given DGS colors for identification (Fig 5) and appropriate scores in the LRT.
Figure 6. Bluefieldius is a sister to Paratarrasius in the LRT. Note the tiny lateral fins and heterocercal tail. A dorsal fin is not preserved in this specimen. The twisted torso not revealing the skull dorsal surface indicates the possibility that the dorsal fin, if present, is still buried in the matrix.
These corrections are improving the scoring of the fish subset of the LRT, which is still ‘in progress’. Perhaps it will always be ‘in progress.’
References Janvier P and Arsenault M 2007. The anatomy of Euphanerops longaevus Woodward, 1900, an anaspid-like jawless vertebrate from the Upper Devonian of Miguasha, Quebec, Canada. Geodiversitas 29 (1) : 143-216. Pradel A, Maisey JG, Tafforeau P, Mapes RH and Mallant J 2014. A Palaeozoic shark with osteichthyan-like branchial arches. Nature 13185. doi:10.1038/nature13195e Sansom RS, Gabbott SE and Purnell MA 2013. Unusual anal fin in a Devonian jawless vertebrate reveals complex origins of paired appendages. Biol Lett. 2013 Jun 23; 9(3): 20130002. doi: 10.1098/rsbl.2013.0002 Zhu Y-A et al (10 co-authors) 2022. The oldest complete jawed vertebrates from the early Silurian of China. Nature 609:954–958. online Woodward AS 1900. On a new ostracoderm fish (Euphanerops longaevus) from the Upper Devonian of Scaumenac Bay, Quebec, Canada. Magazine of Natural History ser. 7, 5: 416-419.
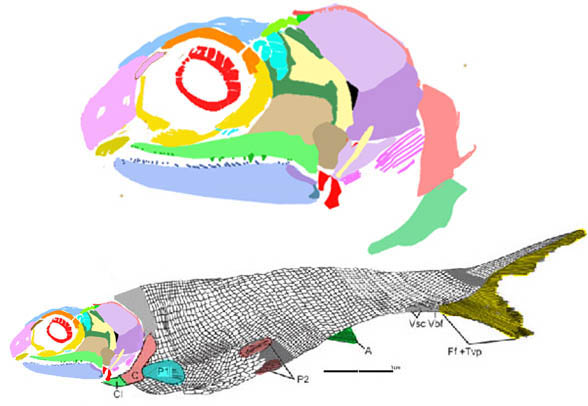
I overlooked this earlier. The mid-sized placoderm Amazichthys (Fig 1) had a full set of dentary teeth, distinct from the placoderm tradition (but see below) and unrelated to similar teeth found in other fish, according to the LRT.
Described in 2022 by Jobbins et al, Amazichthys amazingly preserves impressions (and bone) of the entire fish, but unfortunately the skull is exposed in only a few places in both known specimens. So, maxillary data is currently not available.
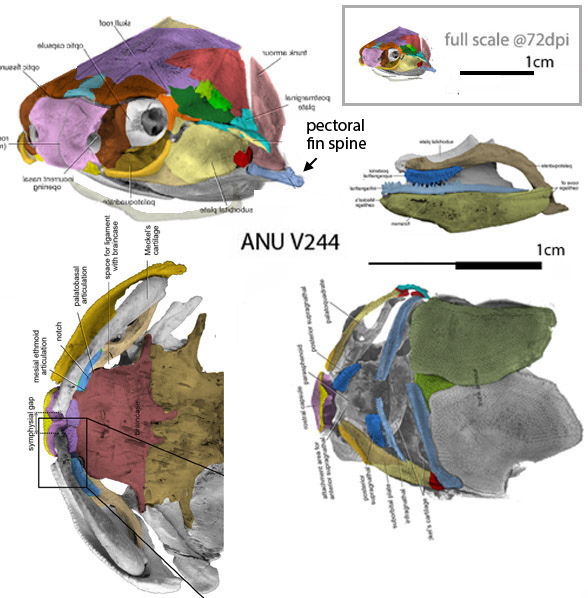
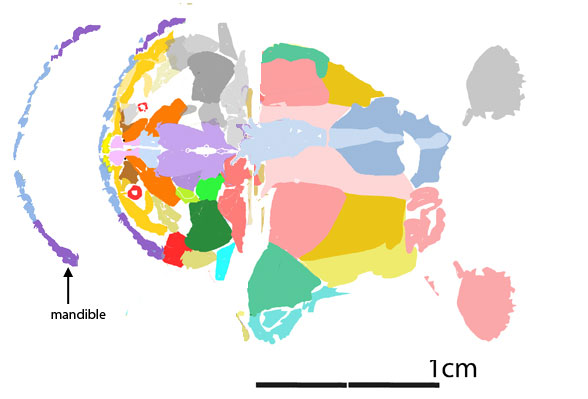
Perhaps no maxilla was present and the dentary teeth worked against the palatine teeth, as in the tiny related unnamed specimen, ANU V244 (Fig 2).
FIgure 1. Amazichthys data. Colors added here. At bottom is the dentary, articular and quadrate. Note the teeth at various angles and shapes.
The authors reported on the mandible, “The bone is rather slender at its biting division and presents a row of at least four, less than 1 mm high, preserved teeth. The jaw is higher in the posterior half, i.e., the bony shaft.”
No other attention was paid to the teeth by Jobbins et al.
Figure 2. The unnamed tiny Late Silurian transitional placoderm, ANU V244 was preserved in perfect 3d. Colors added here. The middle right graphic shows the light blue dentary teeth occluding with the rich blue palatine teeth.
Smith and Johanson reported in 2003, “It is generally thought that teeth are absent in placoderms and that the phylogenetic origin of teeth occurred after the evolution of jaws. However, we now report the presence of tooth rows in more derived placoderms, the arthrodires. New teeth are composed of gnathostome-type dentine and develop at specific locations. Hence, it appears that these placoderm teeth develop and are regulated as in other jawed vertebrates. Because tooth development occurs only in derived forms of placoderms, we suggest that teeth evolved at least twice, through a mechanism of convergent evolution.”
Taking a closer look at a tiny related placoderm, Millerosteus (Fig 1), a carpet of teeth is preserved on the inner dentary. With similar overlapping cheekbones as the ANU V244 specimen (Fig 2) it is likely these lower teeth occluded only with the carpet of palatine teeth. The maxilla was probably absent, as in ANU V244.
Amazichthys trinajsticae (Jobbins et al 2022; Late Devonian) was preserved with a body outline indicating a deep caudal and dorsal fin. Otherwise this placoderm was a larger Millerosteus in most respects, but with a shorter thorax shield. The cheek bones remain unknown and are restored here based on Millerosteus.
References Jobbins M, Rücklin M, Ferrón HG and Klug C 2022. A new selenosteid placoderm from the Late Devonian of the eastern Anti-Atlas (Morocco) with preserved body outline and its ecomorphology. Front. Ecol. Evol. 10:969158. doi: 10.3389/fevo.2022.969158. Smith MM and Johanson Z 2003. Separate evolutionary origins of teeth from evidence in fossil jawed vertebrates. Science 299(5620):1235–1236.
If you ever wondered how the under-slung parasagittal oral elements of jawless fish with a transverse hinge (Fig 1) evolved into the more familiar arc with paired lateral hinges (Figs 2–4), here’s a series of graphics that show a step-by-step evolution. This series applies only to the placoderm clade and all taxa that descended from placoderms.
As recovered earlier, stem tetrapod jaws developed in a different clade of fish. Thus the traditioinal clade, ‘Gnathostomata‘ is no longer monophyletic. See Birkenia for details.
Figure 1. Images from Lanzetti et al 2023, animated here. This is the oral apparatus of Rhinopteraspis after µCT scanning. These are not jaws.
Rhinopteraspis (Fig 1) demonstrates the jawless condition in pre-placoderms. The mobile oral elements opened like a castle draw bridge. Note the transverse hinge of the multiple elements and the parasagittal orientation of the medial elements, unlike the jaws in extant vertebrates.
Figure 2. Qilinyu skull in ventral view with original identifications and revised here. The barely conjoined elements of the mandible that once were the tips of the ancestral oral elements shown in figure 1, now gone. The motion is the same. The axis of rotation has migrated to the lateral skull.
The tentative genesis of jaws in Late Silurian Qilinyu (Fig 2) demonstrates the transition from parasagittal elements in jawless pre-gnathostomes to lateral elements in basal gnathostomes. Note the disappearance of all but the anterior tips of the former mobile oral elements (Fig 1), now tentatively conjoined, but still moving in the same limited arc. This is a very weak jaw.
Figure 3. In Bianchengichthys the jaws are still weak and fragile, but they arc across the margin of the mandible here as the nares rotate from ventral to anterior.
In phylogenetically miniaturized Bianchengichthys (Fig 3) the jaw evolved into a greater arc that matched the rostral margin enabled by reduction of the nasal elements and migration of the once ventral nares. Whether the mandible elements were strongly conjoined or not cannot be determined from this tiny fractured fossil and its gracile mandible. Either way, the mandible was still fragile.
Figure 4 Skull of the small arthrodire placoderm, Coccosteus, demonstrating the evolution of robust mandibles without matching the upper jaw margins. Compare this ventral view with that of Qilinyu in figure 2.
Not all primitive placoderm jaws were gracile and weak. Coccosteus(Fig 4) also had an under-slung jaw, like Qilinyu (Fig 2), but it was sharp and robust. Note the similar ventral narial openings in Coccosteus and Qilinyu. Are these excurrent nares? And why do they not appear in the anterior view of Coccosteus where the ventral nasal plate lacks the perforations seen in ventral view? Here we also see the origin of palatal teeth (blue arc in ventral view) recapitulating in arc and chain of gracile elements the earlier origin of the mandible.
Evolution is a fascinating subject. I’m glad you’re along for the ride.
Figure 5. Bothriolepis oral cavity animated. Here the mandible remains immobile and the nasal becomes mobile, scraping bacterial mat perhaps, or ingesting sand, or scraping coral.
PS Bothriolepis(FIg 5), another placoderm, did not move its mandibles. Instead it moved its nasals, rimmed by scraping premaxillae. The number of mandible elements in Bothriolepis corresponds with those of Rhinopteraspis (Fig 1), including those extra-wide medial elements with a narrow ‘neck’. Based on this example, the premaxilla also evolved several times. And of course it disappeared several times more. That is why phylogenetic analysis can be so vexing to score.


























{kind=link}