It’s SVP abstracts season!!
…and this is number 10 of 10.

Figure 1. Sphaerolepis and Elonichthys to scale. Shown 1.8x lifesize on a 72 dpi monitor. Arrows point to distinct differences.
Unexpected characters in a Pennsylvanian ray-finned fish
Lyons-Weiler, Giles and Friedman (p398)
“the relationships of these early, sometimes morphologically disparate, late Paleozoic rayfins = (Actinopterygii) are uncertain, with little agreement on the temporal origin of major extant clades.”
In the large reptile tree (LRT, 2441 taxa) rayfin fish had several origins.
“With an anatomically generalized postcranium, Sphaerolepis was historically allied to ‘paleoniscoids,’ a paraphyletic grouping of early ray-fins.”
Added to the LRT Sphaerolepis (Fig 1) nests with Elonichthys among the paleoniscids.
“Initial incorporation of Sphaeroelpis [sic] in phylogenetic analyses destabilizes early ray-finned fish interrelationships, generating a large polytomy composed mostly of Paleozoic members.”
In the LRT Sphaerolepis does not destabilize fish interrelationships.
“Sphaeroelpis [sic] emphasizes that many poorly known, apparently generalized, Paleozoic actinopterygians are likely riddled with unexpected characters.”
“likely riddled” is only an issue that needs to be resolved. So…resolve that issue. Science is all about finding out. Add more taxa. Take a closer look. Etc.

Figure 2. Erpetonyx in situ. This is an early diapsid in the LRT, not a bolosaurid.
Computed tomography (CT) data reveals new insights into the evolution of herbivory in bolosaurian reptiles
MacDougall, Ponstein and Fröbisch (p399)
“Primitive bolosaurian taxa, specifically Erpetonyx, exhibit few if any adaptations for herbivory, whereas we observe a cascade of increasing complexity of dental adaptions in more derived taxa, such as Eudibamus, Bolosaurus, and Belebey.”
In the LRT, only Bolosaurus and Belebey are bolosaurs. The other two (Erpetonyx shown in Fig 2) are basal diapsids. Bolosaurs are sisters to diadectids + procolophonids, clades not mentioned by the authors.

Figure 3. Mcnamaraspis, Stenosteus, Cobelodus, Gemuendina and Jagorinia compared in dorsal view and in phylogenetic order. DGS colors added here.
The anatomy of a rhenanid placoderm, Jagorina pandora, revealed by synchrotron radiation microtomography
Pears et al (p482)
“Placoderms (stem-group jawed vertebrates) are central to debates on the evolution of early gnathostome anatomy.”
Not central. The authors don’t realize that jaws developed 3x by convergence in the LRT. Add taxa to find this out for yourself.
“One of the major obstacles to resolving the origin of key gnathostome characters such as jaws, paired appendages, and girdles is the scarcity of preserved endoskeletons, which were only weakly ossified in most placoderms.”
The LRT was able to lump and separate most taxa based on their exoskeletons and/or endoskeletons. All such data is useful.
“Here we reinvestigate the anatomy of Jagorina pandora, a rhenanid placoderm from the Late Devonian of Germany, known for three-dimensionally preserved shoulder girdles, as well as jaws and the branchial skeleton in articulation with the endorcranium. [sic]”
“We provide the first clear three-dimensional description and reconstruction of rhenanid pectoral girdles and fins, revealing previously unknown portions of the dermal and endoskeletal shoulder girdle, which differ significantly from past reconstructions.”
Good to know. In the LRT Jagorina (Fig 3) nests with Gemuendina, Titanicthys, Stenosteus. Cobelodus, Mcnamaraspis and tiny Millerosteus the base of the wider, flatter placoderms.
“We highlight specific similarities between the shoulder girdles of rhenanids and acanthothoracids (see definition below) relative to crown-group gnathostomes, hinting at areas of conflict in existing phylogenies and challenging current theories about the shared ancestral condition of crown-group gnathostome and placoderms.”
Definition: Acanthothoraci (spine chests) is an extinct group of chimaera-like placoderms closely related to the rhenanid placoderms.
Add taxa to relieve those areas of conflict. Gnathostomes had several origins.

Figure 4. Tanystropheus and kin going back to Huehuecuetzpalli.
Caudal autotomy in tanystropheids (Reptilia, Tanysauria): the first known occurrence of tail shedding in archosauromorphs”
Pittinger et al (p494)
“Caudal autotomy (tail shedding) is a defense strategy observed in living salamanders and lepidosaurs.”
Then why do the authors think Tanystropheus (Fig 4) is an archosauromorph? Because someone told them so? Test this yourself to find out.
“The scans provided internal visualization of the fracture plane and associated transverse bony septum; the bony septum extends internally through the centrum, comparable to the autotomous vertebrae of extant lepidosaurs.”
Peters 2007 nested tanystropheids within Lepidosauria. So far, no one has tested this hypothesis, including these authors who insist or believe, without testing that tanystropheids are archosauromorphs.
“along with the transverse caudal septa identified in this study and past reports, indicate that tanystropheids may have been capable of shedding their tails, making this the first known occurrence of this defense mechanism in archosauromorphs.”
Be careful with such phylogenetic ‘firsts’. The authors have the wrong phylogeny and are proudly spreading a myth at a convention. First build your own valid wide-gamut cladogram. Then talk about your focused study.
No examples of severed-tail or tail-less tanystropheids are known.
This fact is not mentioned in the abstract.
Finally, do you see the deep chevrons in the largest Tanystropheus (Fig 4)? Workers have puzzled over these. Note the pterosaur-like attenuated tail, reducing the anchor area for the once massive caudofemoralis muscles. Now imagine those giant chevrons anchoring lower extensions of the caudofemoralis muscles and you have not only powerful leaping muscles but heavy counterweights for that super-long neck AND the third leg of a tripod when Tanystropheus goes vertical.

Figure 5. An undescribed Green River bat to scale with the skull of Microcebus, the dwarf lemur, an ancestor to bats in the LRT. This is full scale on 72dpi monitors
Can the evolution and origins of laryngeal echolocation in bats be revealed by examining cranial growth rate?
Pommery et al. (p499)
“The current bat fossil record does not provide sufficient evidence to determine the origins of laryngeal echolocation.”
Then look beyond the fossil record. The mouse lemur, Microcebus, the smallest living primate, is the ancestor of bats in the LRT.
According towiki/Gray_mouse_lemur ”
“Vocalizations are complex and very high-pitched (ranging from 10 to 36 kHz), sometimes beyond the range of human hearing (0.02 to 20 kHz). These include calls for seeking contact, mating, distant communication, alarm, and distress.”

Figure 6. Leptictis compared to Rhynchocyon and Macroscelides.
The identification of Cretaceous crown placentals is hindered by incomplete fossil records and substantial morphological similarity
Qi et al (p508)
“no unambiguous crown placentals have been confirmed from the Late Cretaceous, although a few candidate taxa have been proposed.”
Not true. Jurassic and later multituberculates are crown placental 2 taxa close to rodents.
“To investigate the reasons behind this discrepancy, we reconstructed ancestral nodes across the mammal tree using 3533 skeletal characters from 46 extant taxa, employing both likelihood-based and parsimony-based methods.”
That’s more characters than necessary (based on experience with the LRT) and too few taxa. And why no fossils? The answer comes in the next sentence of the abstract:
“We found that although Paleocene leptictid (Leptictis dakotensis, Fig 6) is well supported as crown placental closely related to Boreoeutheria, the incompleteness of Late Cretaceous leptictids (Gypsonictops) and Protungutalum fossils poses a major barrier to confidently confirming or rejecting their crown.
Definition: Boreoeutheria is a genomic clade that includes groups as diverse as giraffes, pigs, zebras, rhinoceroses, dogs, cats, rabbits, mice, bats, whales, dolphins, and simians (monkeys and apes). according to Wikipedia.
In the LRT that label for that list of taxa is a synonym for Metatheria = all mammals except monotremes. In the LRT extinct Leptictis (Fig 6) nests at the base of the placental 2 clade and therefore derived from tree shrews and close to sengis like Rhynchocyon (Fig 6), which is ancestral to Phiomicetus and other tenrecs + toothed whales. Add taxa to find this out for yourself.
“This challenge is further compounded by the substantial morphological similarity among ancestors of crown therians, crown placentals, and placental magnorders.
Happy to see that others also view the Mammalia as the most difficult of all clades.
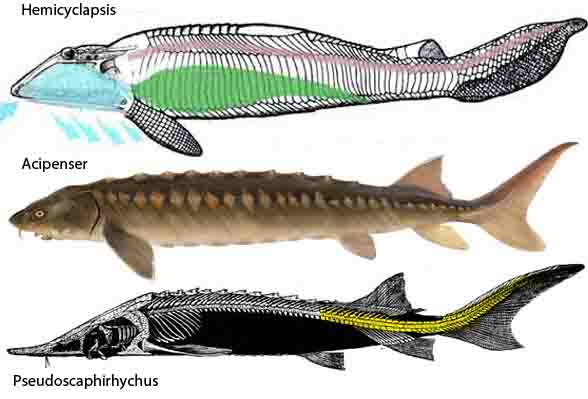
Figure 7. Two extant sturgeons, Acipenser and Pseudoscaphirhynchus, compared to the extinct heterostracan, Hemicyclaspis, a close relative to Zenaspis in figure 4.
A phylogeny for Heterostraci (stem gnathostomes)
Sansom, Randle and Keating (p548)
“Problematic heterostracans with a tessellate headshield (‘tessellate-basal’ model) are often regarded as the plesiomorphic condition for the clade but no phylogenetic analysis has included these taxa.”
The LRT includes only a few of these head-shield taxa, all nesting basal to sturgeons (Fig 7). The authors’ study goes deep, but not deep enough to include sturgeons and kin, and not deep enough to include their ancestor or outgroup from the Cambrian, Haikouichthys.
“Here we review the Heterostraci and present their first comprehensive phylogenetic analysis (131 heterostracan taxa and 55 outgroup taxa). Heterostraci are recovered as monophyletic, and forming a clade together with the Ordovician Pteraspidomorphi near the root of the gnathostome crown group in parsimony analyses.”
The authors are now aware that jaws appeared several time in fish, so there is no such thing as one ‘gnathostome’ clade.
In the LRT Arandaspis is basal to Placodermi (Proaspis is at the base) and Chondrichthyes (Eriptychius is at the base)
“The heavily armored Amphiaspida are recovered as sister taxon to all other Heterostraci. Within the Heterostraci, the Psammosteida are nested within the Pteraspidiformes, which are sister group to the Traquairaspidida, whist Cyathaspidida form a monophyletic group.”
Frankly, those taxa have not been tested in the LRT. There’s still time!
“The results presented here are the first phylogenetic hypotheses of heterostracan relationships and it is hoped a first step into an accurate interpretation of character evolution in this crucial episode of vertebrate evolution.”
Acutally not that crucial… unless sturgeons are your thing.

Figure 8. Rotaryus skull, newly added to the LRT in 11.25. DGS colors added here. Note the distortion.
New insights into the anatomy of the early Permian trematopid Rotaryus gothae (Temnospondyli, Dissorophoidea) using CT data and re-evaluation of its phylogenetic position
Schmitz et al (p554)
“This study reevaluates Rotaryus gothae, a trematopid from the early Permian Bromacker locality in central Germany, using computed tomography (CT) and 3D digital segmentation.”
When added to the LRT Rotaryus (Fig 8) nested with Acheloma and then Platyhistrix in the Dissorophus clade. This hypothesis matches Berman et al 2011.
“The results of these analyses confirm the classification of Rotaryus as a trematopid, but its phylogenetic position varies by dataset and method used.”
Varied? By how much? In which direction?

Figure 9. Origin and evolution of the prepubis in tritosaurs.
Functional morphology of the prepubis in Pterosauria
Schneider, Henderson and Claessens (p555).
“Pterosaurs had a unique pelvic morphology, in which a mobile prepubic skeletal element articulated with an anteroventral facet on the pubis.”
This is incorrect. The prepubis is not unique to pterosaurs (Fig 9). Ancestors had one and the prepubis was not mobile. It had a butt joint with the pubis. The prepubis was a new anchor for sprawling femor adductors as the old femoral abductors, the caudofemoralis muscles anchored on the attenuating tail, became smaller and smaller.
“the homology and function of the pterosaur prepubis remain poorly understood.”
This is incorrect.
First: the prepubis has no homology. It developed de novo as the fenestrasaur tail became attenuated, losing the anchor for the caudofemoralis muscle as fenestrasaurs became bipedal and flapped their forelimbs.
Second: the prepubis also supported the ventral basket of gastralia during bipedal excursions. This became more useful as the skull and wings became larger and larger at the end of a long lever with a fulcrum focused at the acetabulum. You can learn this by building a full scale pterosaur model skeleton.
Third: The anterior extension of the ilia and vertebrae added to the sacrum (Fig 9) are also part of this transition to a bipedal configuration enabling the genesis of flapping.
“An analogous pelvic structure evolved in the Jurassic in Crocodyliformes, in which the pubis articulates with a single facet on the anterior process of the ischium.”
This is incorrect. In crocs the pubis loosens to become mobile and it is attached to a diaphragm-like ventilation structure. It is not an analog.
“To examine the function of the prepubis in pterosaurs, we studied the effects of prepubic movement on body volume and center of mass (CoM) during prepubic rotation and visceral shifting using three-dimensional digital models.”
Note: the authors are ignoring the attenuation of the tail and reduction of the caudofemoralis and the other changes in the pterosaur precursor pelvis (see above) while assuming incorrectly that the prepubis was mobile. Don’t make this same mistake. Look at everything and compare with outgroup taxa.
“We digitally simulated pubic and prepubic movements, respectively, in the extant crocodylian Alligator mississippiensis, in the basal rhamphorhynchoid Rhamphorhynchus muensteri, and in the derived pterodactyloid, Pterodactylus antiquus, using pubic rotation metrics derived from in vivo cineradiographic studies of A. mississippiensis.”
Sounds like they got what they were looking for, but this experiment is ill informed, ill begotten and incorrect in every respect. Redo the experiment to find out for yourself.
“We propose that the pterosaur prepubis served as an accessory breathing mechanism, similar to the respiratory function of the pubis in extant crocodylians.”
One more myth added to the many myths now promoted by pterosaur workers.
The Prepubis of Pterosaurs (and Fenestrasaurs)

Figure 10. Doryaspis specimens in situ. One has an anterior process of the ventral skull, the other does not.
Synchrotron scanning of Doryaspis arctica reveals an exceptionally preserved heterostracan feeding apparatus
Schnetz et al (p556)
“Here, we present the oral anatomy of the Devonian pteraspid heterostracan Doryaspis arctica from the Wood Bay Formation of Spitsbergen, Norway. Doryaspis is known for its unique body plan, with wing-like lateral projections and an extensive ventral process underlying a dorsally located mouth.”
I just became acquainted with Doryaspis today with this photo (Fig 10). Galeaspids have a dorsal immobile oral opening and thus have nothing to do with the origin = genesis of any of the jawed clades. So what does that anterior probe (when present) do? If present on some and not others can we consider it a sign of gender, perhaps used in the process of courtship, seduction and mate choice?
“The oral apparatus of Doryaspis contains a series of pteraspid-like elongated oral plates, arranged transversely across the oral opening, which connect posteriorly to struts, combining to produce a comb-like sieve.”
That comb-like sieve worked like a sewer grating or a kitchen sink drain stopper to keep unwanted material out of the otherwise perpetually open dorsal oral cavity of this bottom dweller (Fig 10) found in groups.
“Our results indicate that at least some pteraspid heterostracans, including Doryaspis,were microphagous suspension feeders, refuting alternative hypotheses suggesting a deposit or predatory feeding style.”
This is incorrect. Other galeaspids were flat, wide, passive bottom feeders, hiding in sand, waiting for prey to walk over their dorsal oral opening. Already buried at the time of death, no wonder fossilization was so perfect.

Figure 11. In the large reptile tree Procolophon nests with Diadectes, and both share a large otic notch, a trait Wiki says makes Diadectes an amphibian. Procolophon is phylogenetically miniaturized = neotonous.
Skeletal accumulations of the parareptile Procolophon trigoniceps reflect fossorial response to Early Triassic climatic instability across southern Gondwana
Smith et al (p585)
“The Lower Triassic parareptile Procolophon trigoniceps is known from hundreds of specimens.”
The LRT does not recover a clade or grade ‘Parareptilia’ Instead Procolophon (Fig 11) is a small (phylogenetically miniaturized) diadectid in the LRT.
“their fossils occur in hyper-abundant skeletal concentrations. lying side-by-side, criss-crossing, or in curled-up poses. In situ cylindrical scratchmarked decline burrow casts occur in the same outcrops.”
That’s interesting.
“The taphonomic evidence supports previous suggestions that P. trigoniceps was a group living, possibly communal, fossorial reptile analogous in its life habits to Gopherus agassizii, an extant North American desert tortoise.”
Sounds like a valid conclusion.

Figure 12. Late Carboniferous Amphibamus reconstructed. DGS colors added here.
Taxonomic diversity and development of Late Carboniferous amphibamiforms from the Mazon Creek Lagerstätte
So et al (p587)
“The earliest anatomically modern representatives of frogs, salamanders, and caecilians all occur in the Triassic; however, there is a significant gap in our understanding of how and when these lineages diversified.”
That gap was resolved in the LRT frogs and salamanders nest together and caecilians nest elsewhere in the Microsauria.
“The numerous larval specimens allow us to identify discrete ossification patterns in Amphibamus life history that establishes a staging table for Amphibamus.”
“Phylogenetic analysis under maximum parsimony and Bayesian inference recovered specimens in derived positions, including FMNH PR 5055 as the immediate sister taxon to Lissamphibia.”
In the LRT Amhibamus (Fig 12) is basal reptilomorph, descended from basalmost microsaurs and lissamphibians – so very close, but on the other side of the ledger.

Figure 13. Small Prolacerta specimen AMNH 9520 from Spiekman 2018 compared to scale with Vellbergia. Scleral rings are labeled scl in the lowest photo.
The paleobiology of Prolacerta broomi: inferences from paleoneurology
Sobral G and Ezcurra MD
“Archosauromorpha is a clade of diapsid reptiles comprising living crocodylians and birds, and all fossil groups more closely related to them than to lepidosaurs.”
The LRT supports and follows this definition except for the diaspid part, which was added to this abstract. Thus in the LRT Archosauromorpha includes living crocodylians and birds, and all fossil groups more closely related to them than to lepidosaurs. That’s how Archosauromorpha also includes Synapsida and more primitive taxa, like Gephyrostegus, back to the Early Carboniferous.
“The group originated in the middle-late Permian, and radiated to a broad array of environments before the origin of the crown, including aquatic and possibly gliding forms.”
In the LRT archosauromorphs, like Gephyrostegus, appeared much earlier, in the Early Carboniferous. Gliders include dermopterans, sugar gliders, ‘flying’ squirrels and gliding multituberculates. Flying members include bats and birds.
“Because no diel (= daily) activity pattern has been formally proposed for Prolacerta, here we analyze the scleral ossicle ring anatomy for the first time based on a new specimen.”
Spiekman 2018 identified a sclerotic ring (Fig 13) in a previous published description of this specimen. Spiekman SNF 2018. A new specimen of Prolacerta broomi from the lower Fremouw Formation (Early Triassic) of Antarctica, its biogeographical implications and a taxonomic revision. Nature Scientific Reports 8:17996 | DOI:10.1038/s41598-018-36499-6
“A phylogenetic flexible discriminant analysis with 96% accuracy confirmed Prolacerta was a photopic (diurnal) taxon,”
This makes sense considering the cold-blooded metabolism.
“This is the first time diel activity patterns have been estimated for stem archosaurs based on the direct measurement of complete scleral ossicle rings”
Good to know!

Figure 14. Subset of the LRT focusing on basal Archosauromorpha including Vaughnictis and Cabarzia nesting at the base of the Protodiapsid-Synapsid split. Note all the large varanopids nest together here in the Synapsida, separate from small varanopids in the Protodiapsida.
Exceptionally preserved skeleton from a lower Permian interdune deposit in southeastern Utah sheds light on early synapsid phylogeny and ecomorphological diversity
Sodano et al (p589)
“The earliest stem mammals diversified during the late Carboniferous and early Permian, with Caseasauria, Ophiacodontidae, Sphenacomorpha, and Varanopidae present in the fossil record by the Carboniferous/Permian boundary ~298.9 Ma.”
In the LRT Caseasauria are not synapsids. Instead Caseasauria nests with Milleretta in the Lepidosauromorpha.
In the LRT Varanopidae were basal synapsids and some derived taxa added a temporal opening to become archosauromorph diapsids (Fig 14), convergent with Lepidosauromroph diapsids. The authors are unaware of this phylogenetic hypothesis due to tradition and taxon exclusion that is minimized by the LRT.

Figure 15. Eudimorphodon premaxilla as identified by Wild 1978. Above, in situ. Below, reconstructed.
Pterosaur phylogeny revisited with novel characters and comprehensive analyses
Song, Jiang and Wang (p590)
“Recent years have witnessed substantial advances in the study of pterosaur phylogeny, marked by the incorporation of an expanding array of characters and operational taxonomic units (OTUs) into different datasets.”
Only more taxa are needed, as shown by the large pterosaur tree (LPT 266 taxa), not more characters (166). Academic cladograms omit large numbers of large and small pterosaurs and their valid outgroups for various in-house reasons.
“Here we assemble a new data matrix covering full taxonomic breadth of Pterosauria, featuring over 550 characters and 170 ingroup taxa.”
This is too few taxa. The LPT includes 251 ingroup taxa. Those extra 81 taxa are al necessary. So are valid outgroup taxa. The LRT gets by with 183 multistate characters. No more are needed.
“Triassic pterosaurs with tricuspid teeth were not found in a monophyletic clade.”
Interesting point. In the LPT the last common ancestor of pterosaurs, Bergamodactylus, had tricuspid teeth. So did pterosaur proximal ancestors, like Cosesaurus, Sharovipteryx and Longisquama (Fig 16). So did Triassic pterosaurs like Austriadraco and Seazzadactylus.
Raeticodactylus and Austriadactylus lacked tricuspid teeth.
Then pterosaurs split between Dimorphodontidae (no tricuspid teeth) and Eudimorphodontidae (only Eudimorphodon (Fig 15) and Arcticodactylus had tricuspid teeth) in the LPT.
More Triassic taxa may someday resolve this issue.
“Anurognathidae was consistently resolved as a basally branching novialoid, forming the sister taxon to a group comprising remaining members of this clade except for Campylognathoides.”
The LPT does not recover the clade Novialoidea. wiki/Novialoidea
Taxon exclusion hobbles this abstract. In the LPT anurognathids descend from similar dimorphodontids. The genus Campylgnathioides (5 tested specimens) gave rise to Rhamphorhynchus (a few dozen tested taxa in the LPT) via phylogenetic miniaturization.
“the earliest diverging clade of Pterodactyloidea”
In the LPT the traditional clade “Pterodactyloidea” is a grade that was achieved 4x by convergence. We’ve known this since Peters 2007.

Figure 16. Longisquama and Cosesaurus compared to the most complete specimen of Mirasaura after tracing elements using DGS colors.
A drepanosauromorph from the Grés á Voltzia fossil Lagerstätte (early Anisian) of northeastern France reveals early diversification of integumentary appendages among Diapsida
Spiekman et al (p593)
This Longisquama-like taxon was published and criticized earlier here: mirasaura-grauvogeli-a-new-fenestrasaur-close-to-longisquama/

Figure 16. Scaphognathus in situ focusing on the dorsal plumes that were identified.
In short: taxon exclusion hobbled this study of high-plumed Mirasaura (named in the published paper, Fig 15). Longisquama has been fenestrasaur (= pre-pterosaur) since 2000 and a lepidosaur since 2007. Drepanosauromorphs are the sister clade to the tanystropheids within Tritosauria in the LRT. The authors are not aware that the diapsid skull architecture evolved twice by convergence, once in Lepidosauria and once in Petrolacosaurus. The authors are not aware that Cosesaurus and certain pterosaurs (Bergamodactylus, Scaphognathus (Fig 16) and Jeholopterus) also had dorsal plumes. Finally, the Mirasaura skull (Fig 14) is more like that of a Cretaceous cosesaur, Occuludentavis, a taxon omitted from the abstract and paper.

Figure 17. Basal crocs. Decuriasuchus and Gracilisuchus are found in both croc and dino lineages.
Combining perspectives on archosaur ontogeny reveals the first multitaxic assemblage of “sphenosuchians” from the same Upper Triassic vertebrate community
Srivastava et al (p594)
“The earliest-diverging crocodylomorphs comprise the paraphyletic “Sphenosuchia” (i.e., non-crocodyliform crocodylomorphs). “Sphenosuchians” represent the first pulse of crocodylomorph evolution in the Late Triassic, but the “sphenosuchian” record is sporadic, with most taxa diagnosed from one specimen,”
In the LRT the earliest-diverging crocodylomorphs are all bipeds (Fig 17). These include Scleromochlus and Lewisuchus. Quadrupedal Sphenosuchia are more derived.

Figure 18. Torosaurus skull in dorsal view.
Postcranial anatomy of Torosaurus (Ceratopsidae: Chasmosaurine) from the Upper Cretaceous (Maastrichtian) Ojo Alamo Formation of the southeastern San Juan Basin, New Mexico, USA
Straley, Loewen and Srtich (p600)
“We document the first, relatively complete postcranial skeleton of a large individual referred confidently to Torosaurus (EMK 0001) from the Maastrichtian Ojo Alamo Formation”
Great!

Figure 19. Eschrichtius-robustus, the gray whale is the most basal mysticete tested in the LRT with a skull similar to Desmotylus and Beheomotops.
Can suction feeding be determined for fossil cetaceans? Morphological criteria for identifying suction specialists
Strauch et al (p601)
“Most secondarily aquatic mammals employ suction to some extent in the feeding process (e.g., transporting grasped prey to the back of the oral cavity), with many converging on feeding strategies that use suction for prey capture.”
Transporting grasped prey to the back of the oral cavity is usually what the tongue does. Suction feeding is different. Suction involves rapidly expanding the buccal cavity to produce an inrush of water + prey in frogfish and blue whale manner.
I wish the headline was not in the form of a question. Make your headline a statement.
“Suction feeding has repeatedly evolved in odontocetes, with notable examples in extant sperm whales and beaked whales.”
Those taxa don’t rapidly expand the buccal cavity. Instead they have teeth for biting and a tongue for pulling and tasting.
“Here, we aim to develop a scoring system that (1) quantifies specialized suction feeding morphologies from a suite of morphological characters and (2) predicts suction feeding in fossil whales.”
This isn’t going to end well.
“Our results suggest that weak jaw adductors, a robust tongue (or hyoid), and dental wear are the strongest predictors of suction feeding.”
And yet rorquals have no teeth. Neither do frogfish. “although the sperm whale’s tongue is rather large (about one m long in adult males; Tomilin, 1967), relative to the tongues of other odontocetes, it is exceptionally short, wide, and thick.”

Figure 20. Pholidogaster skulls compared to Colosteus and Osteolepis. Panchen reconstruction on left includes a premaxilla that is too wide. At right revised width to fit premaxilla tracing, pectoral girdle and in situ lacrimal and cheek bones.
Evaluating trends in mandibular form and function across vertebrate water–land
transitions
Strong and Pierce (p603)
“As one of the most pivotal events in vertebrate evolution, the origin of tetrapods ~390 Ma involved major changes throughout the body plan as these animals first became adapted to lifeon land.”
This is incorrect. The origin of limbs from fins was not pivotal and did not involve land, but fingers ultimately made that possible. The only noticeable differences between Panderichthys and Trypanognathus are a smaller overall size, a shorter skull (neotony) and four fingers where fin rays once were. Adaption to life on land was a long transition, not a pivotal event.
“One of the most prominent modifications involved the feeding apparatus: whereas aquatic vertebrates often employ suction-based mechanisms to capture and ingest food, the different fluid properties of water versus air preclude this technique on land.”
The authors are jumping the gun: assuming that fingers = land dwellers. Fingers developed for underwater passage through thick vegetation and/or shallow beaches. The authors are also assuming suction-feeding in trap-like pre-tetrapods.
“Here, we test the long-standing notion of a core trade-off between suction-feeding versus biting, Our results show that extant clades exhibit phylogenetically structured morphofunctional solutions to the challenge of feeding in different environments and that these are distinct from the trends exhibited by extinct taxa, highlighting the need for caution when relying on mandibles for deeptime reconstructions of feeding mode.”
The authors concluded: “highlighting the need for caution”

Figure 21. Hemiprotosuhus image from Desojo and Ezccura 2016. Colors added. This taxon is derived from Ticinosuchus, basal to aetosaurs.
Early development of aetosaurs (Archosauria, Pseudosuchia)—a key to their success in the Late Triassic ecosystems?
Talanda et al (p 612)
“Here we calculated the prevalence of various omnivore/herbivore vertebrates in the late Carnian and the entire Norian in all geological units that yield any of them. We found that aetosaurs (Pseudosuchia) were present in a significant majority (79%) of them, outnumbering other groups like prosauropods.”
That’s interesting. Strength in numbers.
“Here we present a nearly complete skeleton of a very young aetosaur from Wozniki (Poland).”
In the LRT Middle Triassic Ticinosuchus (Fig 21) is a basal aetosaur.

Figure 22. Gnathosaurus skull with standard DGS colors.
A remarkable new 3D cranium of the ctenochasmatid Gnathosaurus reveals new anatomical details of the genus
Theda et al (p615)
“Here, we present a new, well-preserved Gnathosaurus cranium from the Painten quarry. It is positioned upright on a limestone slab with its palatal side buried in matrix. It is three-dimensionally preserved, showing almost no signs of crushing or distortion, and the only missing part is the rostrum tip.”
“the presence of a premaxillary crest here indicates it had reached, or was close to, sexual maturity.”
A crest was identified earlier here (Fig 22) in 2021.
“The new cranium reveals novel anatomical details of Gnathosaurus, such as an ascending ramus of the maxilla, giving the anterior end of the nasoantorbital fenestra a slitlike appearance.”
That’s the naris – or what’s left of it.
“Both, the already known and the new cranium, possess a similar lacrimal, consisting of a posteriorly curving jugal process and a marked, horizontal supraorbital crest. Although present in some other pterosaurs, this structure is large here and has not specifically been discussed before.”
Those elements have been illustrated before: And see figure 22.

Figure 23. Sisters to Microtuban include No. 42 (more primitive) and Jidapterus (more derived).
Giant size convergently evolved multiple times in Azhdarchidae (Pterosauria, Azhdarchoidea)
Thomas and McDavid (p 618)
“Azhdarchoidea is a clade of pterosaurs which includes the largest-ever flying animals.”
This is incorrect twice. First: Azhdarchids were flightless with wings too small for the rest of their morphology. The distal wing phalanges were vestiges. As in the biggest flightless birds, first the bird became flightless. That enabled them to then become giants. Second: Traditional azhdarchoids include azhdarchids and unrelated tapejarids, which had large wings and were volant.
“The evolutionary history and interrelationships of this clade remains incompletely understood and debated.”
Peters 2007 split azhdarchids from tapejards. The large pterosaur tree (LPT) includes a long list of pre-azhdarchids arising from a tiny unnnamed dorygnathid. The LPT includes a long list of pre-tapejarids.
“We combine multiple preexisting phylogenetic datasets with 23 new OTUs (= taxa) and 51 new characters, resulting in the most comprehensive phylogenetic analysis of pterosaurs to date, with a focus on Azhdarchoidea.”
If they are combining prior studies, then they are omitting pre-azhdarchid and pre-tapejarid taxa. So this study is doomed by following tradition, rather than adding taxa.
“Outside of Azhdarchoidea, Rhamphorhynchinae and Wukongopteridae are resolved as monophyletic, rather than as grades on the lineage leading to Pterodactyloidea.”
The authors are unaware (due to taxon exclusion) that Pterodactyloidea has been a grade, not a clade, since Peters 2007.
“Inabtanin and Microtuban are recovered as basal azhdarchomorphs.”
In the LPT Microtuban (known from only a wing, Fig 23) is a proximal taxon outside the Azhdarchidae along with SOS2428 and SOS2179. Inabatanin is known from jaw tips, and part to most of the wing, so not tested in the LPT.
“The oldest unambiguous azhdarchid material is early Cenomanian, but this material is deeply nested within the clade, suggesting a cryptic radiation of azhdarchids in the late Early Cretaceous.”
Not sure where the authors want to draw the line, but the Middle Jurassic dorygnathid Dearc is the basalmost taxon in the azhdarchid clade here: http://reptileevolution.com/MPUM6009-3.htm
“Gigantism evolved independently four times in Azhdarchidae: Cryodrakon, Hatzegopteryx sensu lato, Arambourgiania philadelphiae, and the Q. northropi-Thanatosdrakon clade.”
In the LPT big-winged Thanatosdrakon (Fig 24) is the largest Pteranodon, so not related to azhdarchids. The rest of the taxa are known by scraps = a few bones.

Figure 24. Thanatosdrakon compared to Quetzalcoatlus showing the distinct morphologies.
PerissOrigin: re-examining the phylogeny of early perissodactyls reveals unnoticed dispersals of the first North American, European, and Asian ‘horses’
Tissier and Smith (p618)
“Among those early genera, Hyracotherium, one of the earliest ‘horses’, was considered as the most emblematic, and the most diverse in terms of species.”
In the LRT Hyracotherium is basal to rhinos and tapirs, not horses – but VERY CLOSE as a sister to Eohippus and the rest of the horses.
“We used a variety of outgroups, including Arctocyonidae, Phenacodontidae, Radinskya, and Cambaytheriidae.”
None of these cherry-picked taxa are perissodactyls in the LRT. Dorcatherium and Theosodon are outgroup taxa to the clade Perissodactyla.

Figure 25. At left, Late Paleocene, Plesiadapis. At right, Castor the extant beaver to scale. DGS colors added here.
Reinvestigation of the auditory bulla and postcranium of the Plesiadapis genus and itsimplications for the phylogenetic affinities of plesiadapiforms
Toussaint, Godinot and Ladveze (p622)
“Although their paleoecology and functional adaptations are increasingly well understood, their phylogenetic relationships within Euarchonta, particularly with Euprimates, remain debated.”
In the LRT rodent-like plesiadapiformes (Fig 25) are related to multituberculates and rodents, not primates. Porcupines and beavers (Fig 25) are similar.
“Recent cladistic analyses have supported the Euprimateformes hypothesis,”
Not the LRT. Far from it. In the LRT Primates had a separate origin closer to carnivores among the placnetal 1 taxa. Primates never had oversized gnawing incisors and a diastema behind them.
“Our analyses reveal that the auditory bulla of Plesiadapis is not petrosal-derived and that its first pedal digit bears a claw, not a nail.”
As in rodents and multituberculates, not primates.
“Incorporating these new data into the most recent character matrix for early euarchontan relationships does not support the Euprimateformes hypothesis, but instead a sister group relationship of Plesiadapiformes and Euprimates.”
Only taxon exclusion produces such odd = untenable = dissimilar interrelationships. Add taxa. Repeat the LRT where all sister taxa look like one another as they model microevolutionary events.

Figure 26. Polypterus is a lungfish relative
The earliest fossil evidence of stem Polypterids
Tumelty, Giles and Igielman (p627)
“Actinopterygians diversified significantly in the early Carboniferous following the end-Devonian mass extinction.”
In the LRT there is no monophyletic clade Actinopterygia. Rather several clades converged on a ray-fin morphology after a Cambrian split.
“The earliest diverging group of actinopterygians are the Polypteriformes, but their fossil record only stems back to the Triassic, despite molecular clock divergence estimates predicting a late Devonian-early Carboniferous origin.”
In the LRT Polypterus (Fig 26) and kin are close to lungfish, so their origin predates the Devonian, and they are far from the earliest diverging group.

Figure 27. Click to enlarge. The origin of the pterosaur wing and whatever became of manual digit 5?
A new structural model for the flight patagia of pterosaurs
Unwin et al (p632)
“The current model, primarily founded on exceptionally well-preserved examples of Rhamphorhynchus, has been widely, though not universally, accepted for the last two decades but reassessment of the fossil evidence shows that the model is flawed in several respects”
‘The current model’ is defined later in the abstract where the authors discuss the internal structures, not the planform of the wing. Unwin promotes the batwing model with a deep chord wing membrane attached to the ankles and a uropatagium binding the hind limbs and lateral toes based on a mistaken interpretations of Sordes.
“Natural and UV light photography and microscopy was used to compile data from 80+ specimens of pterosaurs from the Upper Jurassic Solnhofen Limestones of Germany in which wing membranes are preserved, including examples of Anurognathus, Rhamphorhynchus, Scaphognathus, Pterodactylus, Aurorazhdarcho, and Ctenochasma.”
Good taxon list. Where is Sordes? It also needs the UV treatment.
“This new model includes elements of older models but is distinguished by several unique features, most notably the aktinofibril-elastic fibre network.” Elastic fibres (previously interpreted as muscle fibres but lacking diagnostic features of these tissues),”
“This dynamic structure, and its consequences for the mechanical properties and performance of the wing membranes, was fundamentally different from the functionally passive model based on aktinofibril dominated wings.”
Figure 28. Tanystropheus and kin going back to Huehuecuetzpalli.
A Middle Triassic marine tanysaurian with soft tissue preservation from Yunnan, China
Wang et al (p648)
“diverse non-archosauriform archosauromorphs, including the enigmatic allokotosaurs, the beaked rhynchosaurs, and the polyphyletic “protorosaurs.”
In the LRT the first two are lepidosauromorphs. Protorosaurs are archosauromorphs.
“Recent phylogenetic studies have clarified the relationships among “protorosaurs,” establishing Tanysauria as a monophyletic group representing one of the earliest diverging archosauromorph lineages.”
“The clade Tanysauria was described by Stephan Spiekman and co-authors in 2024.”
According to Spiekman et al., the clade Tanysauria includes all taxa that are more closely related to Tanystropheus longobardicus, Dinocephalosaurus orientalis, and Trachelosaurus fischeri than to Protorosaurus speneri, Prolacerta broomi, Mesosuchus browni, Azendohsaurus madagaskarensis, or Proterosuchus fergusi.[1]
In the LRT Tanystropheus (Fig 27) and Macrocnemus (Fig 27) are lepidosauromorphs close to Huehuecuetzpalli (Fig 27), a taxon omitted from other tanystropheid cladograms. Meawhile Protorosaurus + Prolacerta + Proterosuchus are convergent archosauromorphs. So the clade Tanysauria, as defined, is a junior synonym for Lepidosauromorpha in the LRT.

Figure 29. Toothless Early Cretaceous Archaeorhynchus is the last common ancestor of all extant birds in the LRT.
The evolution of palatal development in the common ancestor of living birds
Wilson et al (p663)
“The earliest divergence of crown birds produced two lineages, the Pan-Palaeognathae (‘ratites’ and their kin) and Pan-Neognathae (all other birds).”
The LRT confirms this. Early Cretaceous Archaeorhynchus (Fig 29) is the last common ancestor of extant birds tested in the LRT. Archaeorhynchus is not mentioned in the abstract.
“Surprisingly, a recently discovered Cretaceous toothed bird, Janavis finaldens, was found to possess a pterygoid which is remarkably similar to those of extant members of Galloanserae (fowl).”
The authors are pulling a Larry Martin. Don’t rely on one bone, like a pterygoid. Test the whole animal! Or the whole skeleton, if that’s all you have.
Janavis is known from scraps. Among the few skull bones is a complete pterygoid that is not a match for chickens and ducks. If it is an ichthyornithiform, then that represents a sterile branch in the LRT. Yanornis nests at the base of the ichthyornithiformes in the LRT. Gansus + Honshanornis are next. Then four other nodes… then Archaeorhynchus, the last common ancestor (LCA) of birds.
With that we have come to the end
of the batch of SVP 2025 abstracts of interest. I learned a few things. Hope you did, too.
See you next year with a new batch of SVP abstracts.















