
You might have noticed
the first dorsal fin is the only fin not lobed in Latimeria, the coelocanth (Figs. 1, 2). The focus on lobe fin fish has always been on the fleshy pectoral and pelvic fins — the ones that universities teach are evolving toward arms and legs. Coelacanths sometimes use these lobed fins in an alternating paddle-like fashion, almost as if they are walking, so that fits that scenario.
The problem is,
extant coelocanths prefer the depths and never even thdeink about crossing land or beaching themselves, or fingering their way through weed-choked swamps. So why did coelocanths have lobed fins at all if they were not using them in a fin-to-limb scenario as textbooks teach? And what about those other fins: the parasagittal tail tuft, second dorsal fin and anal fin? Why would these also have fleshy lobes?

As I assemble this post, at this point I have no answer.
A little research is needed. So… moving forward…

Let’s start with a phylogenetic context.
In the large reptile tree (LRT, 1890+ taxa, subset Fig. 3) lobefins arise from a series of spiny sharks, like Diplacanthus (Figs. 2, 4) near the base of the bony fish clade. Prior to spiny sharks is a clade of Osteoglossiformes and Pachycormiformes, which have robust fins created only by tightly-packed rays without membrane between them. So clades were experimenting and our answer could be no more complex than that. Still, if there is an inherent advantage to fleshy lobes, wouldn’t you like to know what it is? According to the LRT (subset Fig. 3), fleshy lobes evolved several times by convergence, all in the same major clade of bony fish that led to tetrapods.
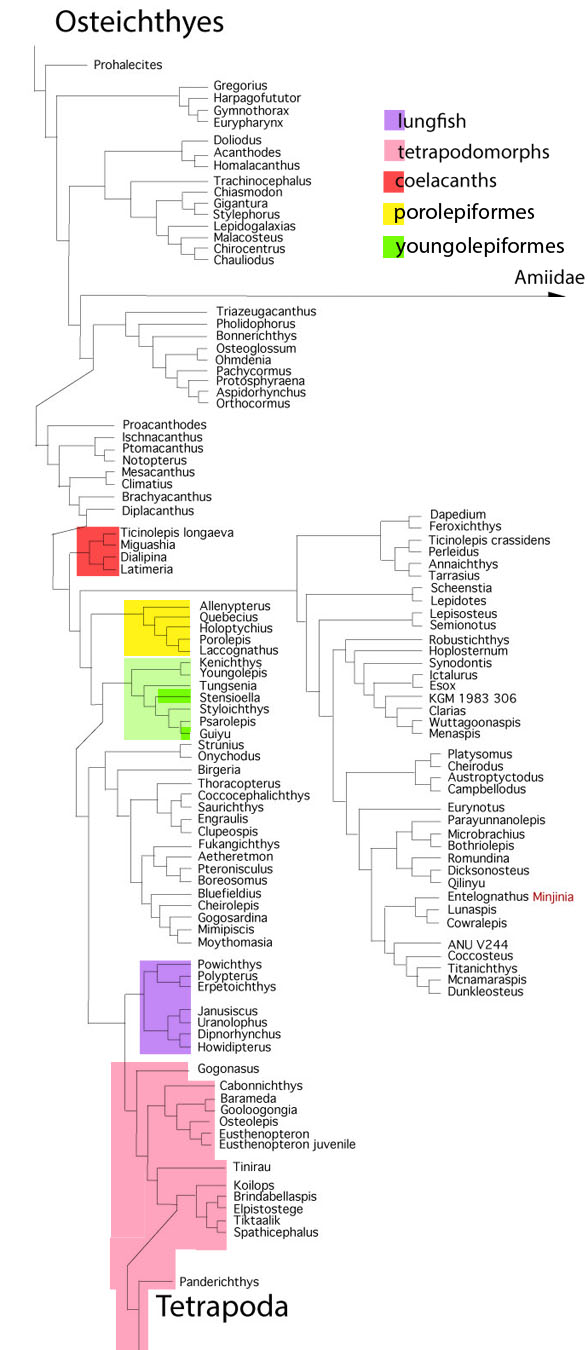
Lobefin fish don’t all nest together
in the LRT (Fig. 3). Not all lobefin taxa have four, let alone six lobe fins (Figs. 2,4). Some don’t have any lobes. They have ray fins instead. This is evolution at its most intriguing and not what they teach in university textbooks and lectures. This (Fig. 3) is what you recover when you add taxa and let the software decide without a priori “Pulling a Larry Martin“.

Putting our focus back on coelocanths
(Fig. 2) late-surviving, but primitive Middle Triassic Ticinolepis had tightly bound pachycormiform-type rays without webbing.
One Middle Devonian coelacanth appears to be instructive.
Miguashaia (Fig. 2) has lobes, but no bones in the lobes. Instead it has two sets of rays per fin at the periphery of the empty lobes. A short inflexible proximal set of rays acts as a base for a long flexible distal set of rays. Latimeria lobes are filled with robust bones, but lack the short inflexible proximal set of rays found in Miguashaia. My guess is Miguashaia had cartilaginous internal precursors to bones to support the otherwise empty lobes and their fan of short and long fin rays.
This pattern is repeated in porolepiformes
(Fig. 4) in parallel by convergence.

There are several studies on fat storage in Latimeria.
None of the fins are used vigorously. In the depths, a slow-motion lifestyle is standard and the coelocanth is no exception. If meals are infrequent, fat storage and its affect on buoyancy can be critical. Latimeria swims like Balistes, the queen trigger fish, lazily undulating its lobed dorsal fin and matching lobed anal fin (Fig. 5). So that explains the muscular lobes in those fins. The big tail is only used for maximum acceleration during prey capture and in escape.
According in explorer, Han Fricke, talking in Wired.com about coelocanths:
“They have the slowest metabolic rate known among vertebrates. We made a calculation that a coelacanth needs, for its resting metabolism, 3.8 milliliters of oxygen per kilogram per hour. A tuna needs 400 milliliters. Because coelacanths are always burning at a low metabolic flame, they are able to live in low-energy areas, where there isn’t much food. The lava fields are a low-product habitat. They need about 12 grams of food a day. This is probably the secret of their evolutionary success. They live where hyperactive fish cannot survive.”
That still doesn’t explain the fleshy pectoral and pelvic fins
in Latimeria. Coelacanths are nocturnal and spend their days sleeping in caves. “They move so slowly.” JLB Smith said “this gives you impression they crawl on their fins at the bottom of the sea, but they don’t. They don’t even touch with their fins.” Images of resting coelacanths show the pectoral fins folded alongside, not stretched out laterally for support, as in flatter, bottom-hugging lungfish and tetrapodomorphs.

On those rare occasions
when massive Latimeria accelerates with every ounce of its being to capture prey in pitch dark, a moment later it will need to brake to a stop or risk crashing into the underwater sea cliffs in its neighborhood. Perhaps that’s where muscular pectoral and pelvic fins come into play, extending laterally like air brakes on a Navy jet (Fig. 6), and able to take the occasional bump and jolt without crippling injury. So that might explain the muscular pectoral and pelvic fins in coelacanths.
Getting back to phylogeny, and niche partitioning…
Coelacanths became adapted to a deep sea niche, beyond light. They lose the maxilla and find prey via electrorerception while retaining a taller than wide body shape (the best shape for fast dashes). Cupello et al. 2019 note, “Fossil coelacanths are registered in shallow brackish, fresh or marine waters, while extant coelacanths inhabit moderate deep marine environments.”
By contrast,
closely related porolepifomes (Fig. 3 in yellow) evolved an increasingly wider than tall body shape as they became bottom feeders in shallow sunlit waters, like anglers, catfish and tetrapodomorphs. So at this node bony fish were finding a wide variety of separate niches in the Silurian and Devonian.
Given that lobe fins can appear here and disappear there,
the original impetus for lobefins remains obscure, other than to say this clade of bony fish were labile, experimenting with several types of fins, a variety of body cross-sections, preferred water depth, activity levels, etc. It is noteworthy that increasing size increases the size of the lobes in coelacanths. Smaller basal taxa in all lobefin clades don’t have such large lobes in their fins, if they have them at all.
This scenario for the appearance of large lobe fins in deep sea Latimeria
seems to make sense and is based on a phylogenetc context (Fig. 3). The only fin not discussed above is the tail tuft. Hissmann and Fricke 1996 covered that, describing how the tiny epicaudal fin is used “in precise maneuvering in the fish’s native, structurally complex habitat. A possible sensory function is also advanced.”
PS.
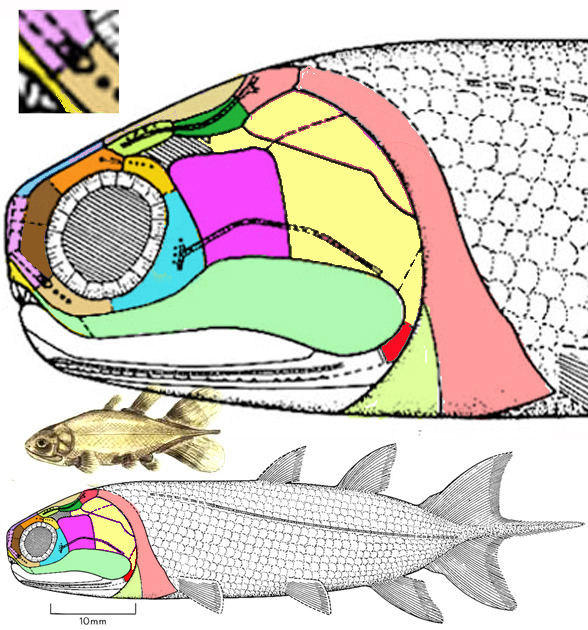
Strunius (Fig. 7) is traditionally considered a lobefin fish despite lacking lobe fins. This identification is based on skull bones and a centrally lobed caudal fin. Basically this a tiny coelacanth that stayed in the shallows and retained its maxilla (light green jaw bone). With so much less mass, acceleration and braking was that much easier and it could see where it was going in the shallows. In the LRT (Fig. 3) Strunius nests at the base of a clade that lacks lobe fins and includes the small, fast, open sea, schooling anchovy, Engraulis, which developed convergent with other ray fin fish on the other major branch of bony fish evolution (not shown in figure 3, so see the LRT).
References
Cupello C et al. (5 co-authors) 2019. The long-time adaptation of coelacanths to moderate deep water: reviewing the evidences. Bull. Kitakyushu Mus. Nat. Hist. Hum. Hist., Ser. A, 17: 29–35.
Hissmann K and Fricke H 1996. Movements of the epicaudal fin in coelacanths. Copeia 3(1):606–615.
Miyake T et al. (15 co-authors) 2016. The pectoral fin muscles of the coelacanth Latimeria chalumnae: Functional and evolutionary implications for the fin-to-limb transition and subsequent evolution of tetrapods. Anatomical Record (Hoboken) 299(9):1203–1223.
wiki/Dialipina
wiki/Latimeria
wiki/Miguashaia
wiki/Actinistia
wiki/Coelocanth
