
Shigeno et al. 2008 write:
“Cephalopod head parts are among the most complex occurring in all invertebrates. Hypotheses for the evolutionary process require a drastic body-plan transition in relation to the life-style changes from benthos to active nekton.”
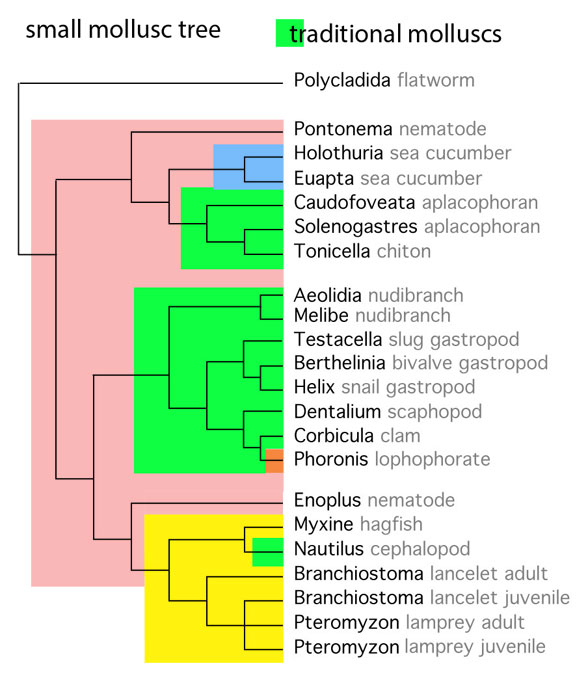
Shigeno et al is following the traditional model (Fig. 1), that the clade Mollusca was monophyletic and the last common ancestor of all mollusks would also be a mollusk. The revised model (posted yesterday) posits a polyphyletic Mollusca, with aplacophorans, snails and cephalopods having three separate ancestries, the latter within basal chordata (e.g. lancelets and hagfish), taxa not considered as outgroups by prior workers.

Shigeno et al. 2008 write and cite:
“How the cephalopod head and associated structures evolved remains largely uncertain
(Packard, 1966, 1972; O’Dor and Webber, 1986).
Unfortunately, Shigeno et al. did not perform a phylogenetic analysis. Instead the authors followed their textbooks. Unfortunately, so did the cited authors.
Shigeno et al. 2010 wrote:
“Ever since this pre-Darwinian academic debate, many zoologists have indulged in a long lasting discussion of how the cephalopod body plan and their organ systems can be linked to those of vertebrates (e.g. Packard, 1972; O’Dor and Webber, 1986). To address the linkage between these two phylogenetically distant taxa, data to infer the original condition was required.”
No prior authors went to the trouble of creating their own phylogenetic analysis, but noted comparisons and indulged in conjecture. No one questioned the monophyly of traditional mollusks.
Shigeno et al. 2008 write:
“Determining these transitions, however, has been elusive because of scarcity of fossil records of soft tissues and lack of some of the early developmental stages of the basal species.”
Taxon exclusion has been the real problem. No one compared basal chordates (and echinoderms) to mollusks in analysis. That’s always the first step. Otherwise you’re on shaky ground. Better to just add taxa to minimize taxon exclusion.
“Here we report the first embryological evidence in the nautiloid cephalopod Nautilus pompilius for the morphological development of the head complex by a unique assembly of multiple archetypical molluscan body parts. Using a specialized aquarium system, we successfully obtained a series of developmental stages that enabled us to test previous controversial scenarios.”
In 2008 they could have added a few taxa. The solution for cephalopod origins did not have to wait an extra 13 years.
By the way, lancelet, hagfish and lamprey hatchlings do not go through a stage in which they are fed by a Nautilus-like yolk sac (Fig. 2) for a year prior to hatching. Instead those chordates follow the ancestral nematode-like strategy and develop from a gastrula more directly to an elongate shape. That many derived fish also develop a Nautilus-like yolk sac as embryos is another example of convergence. The Nautilus embryo goes through a metamorphosis while developing in which the once posterior anus and funnel rotate ventrally until they are beneath the mouth, rotating the dorsal mantle to the aboral (= furthest from the mouth) side of the hatchling (Fig 3).

Shigeno et al. 2008 write:
“Our results demonstrate that the embryonic organs exhibit body plans that are primarily bilateral and antero-posteriorly elongated at stereotyped positions.”
Bilateral and antero-posteriorly elongated = chordate body plan.
“The distinct cephalic compartment, foot, brain cords, mantle, and shell resemble the body plans of monoplacophorans and basal gastropods.”
Their ‘foot’ = buccal cirri or tentacles. That is not correct.
Gastropods have a single mass for a foot posterior to the mouth. A better match for Nautilus tentacles is found in basal chordates where the buccal cirri are also finely divided and surrounding the mouth.
Gastropods have eye spots at the tips of tentacle-like structures. Cephalopods and chordates have internal lateral eyes, outgrowths of the brain and hood.
Nautilus has a shell and chordates do not. In the present hypothesis the shell arises after the 600 million-year-old split from chordate ancestors. The siphuncle that drills through each septa in Nautilus is a remnant of the hagfish notochord buried beneath the developing shield and mantle in the hatchling Nautilus (Fig. 3). Chordates swim with lateral undulations. This is lost in cephalopods in which the former posterior anus and funnel rotates 180º so that the funnel produces ‘backwards’ jet propulsion.

Shigeno et al. 2008 write:
“The numerous digital tentacles of Nautilus develop from simple serial and spatially-patterned bud-like anlagen along the anterior-posterior axis, indicating that origins of digital tentacles or arms of all other cephalopods develop not from the head but from the foot.”
That might be true if gastropods were ancestral and the clade Mollusca was monophyletic. However, the small mollusk tree (Fig. 4) nests cephalopods with chordates, so another hypothetical scenario needs to be considered. A valid phylogenetic context is vital before any other testing or conjecturing is initiated.
“In middle and late embryos, the primary body plans largely change to those of juveniles or adults, and finally form a “head” complex assembled by anlagen of the foot, cephalic hood, collar, hyponome (funnel), and the foot-derived epidermal covers.” [analagen = primordium or embryonic structure subject to change during ontogeny]
This is no longer possible when chordates are added to the taxon list. Nautilus demonstrates the U-turn of the once posterior anus (and funnel) to the anterior, but below the cirri and mouth, not above as in gastropods or to the right body wall, as in sea slugs. Note the funnel is a muscular organ designed to forcefully contract and expel a jet of water. Gastropods don’t have that. Basal chordates do have a primordial funnel in the form of the atrium that also surrounds the gills, but has a separate opening from the anus. The funnel and anus merge in Nautilus when the long straight shell develops in the Ordovician, then the coils develop in later taxa.
“We suggest that extensions of the collar-funnel compartment and free epidermal folds derived from multiple topological foot regions may play an important role in forming the head complex, which is thought to be an important feature during the body plan transition.”
The authors say, “we suggest” and “may play” because they don’t have evidence. They are guessing because they don’t have a valid phylogenetic context. They are relying on textbooks and traditions that never tested chordates and cephalopods together. Yesterday’s phylogenetic analysis (Fig. 4) separate cephalopods + chordates from gastropods and from aplacophorans + polyplacophorans (= chitons).
Summary: The case for a cephalopod – chordate connection is stronger
(= more parsimonious) than the traditional cephalopod – gastropod connection. This extends to the embryology of Nautilus.
Postscript:
Andrew Packard 1966 listed 13 ways in which fish and coleoids (= cephalopods without shells) resembled one another in niche, predatory behaviour, dynamic interaction, etc. Packard did not discuss basal chordates, nor did he perform a phylogenetic analysis. So, while his list is insightful it is also full of conjecture.
From the Packard 1966 abstract (actually more of a list):
“Resemblances between cephalopods and fish:
1. Modern cephalopods (coleoids < 1000 species) resemble modern fish (30,000 species) more closely than any of their ancestors did. They have not been replaced by the more diversified group in geological time.
2. The main body of the article (pp. 245–283) reviews these resemblances. They are to be found at all levels of analysis.
3. Basic physiological mechanisms of molluscs (pp. 262–5) have been incorporated into systems with performances comparable to those of vertebrate systems. For instance the cephalopod locomotory system (pp. 249–56) and hydrostatic control system (pp. 256–60), structurally very different from their fish counterparts, have similar adaptive radiation.
4. Behaviour (p. 278) and growth of the brain (p. 265) are characteristically vertebrate-like.
5. Cephalopods and fish are considered as occupying the same broad adaptive zone though modal differences (pp. 283–5)-in reproductive habits, growth rate and light-dependent behaviour where extraocular photoreceptors appear to be important – mean that they occupy different areas within that zone.(ii) Evolution of convergence
6. Evidence is presented (pp. 287–293) for considering the convergence as due not merely to similar physical demands of the marine environment, but to dynamic interactions between cephalopods and vertebrates from the late Palaeozoic onwards.
7. The convergence was set on its way when the two groups, independently of each other, acquired locomotory methods that allowed them to increase in size.
8. It is argued that reduction and eventually complete loss of the chambered shell (in all but sepiids) was an evolutionary response to the needs of increased mobility and to the need to go deeper as vertebrate predators pushed out into oceanic waters.
9. The ammonites (pp. 291–2) present a partial model of the course that coleoid ancestors may have taken.
10. Coleoids subsequently reinvaded surface and coastal waters, competing successfully in a teleost habitat. Their most striking adaptations are ones that they share with teleosts.
11. Behavioural interactions in a vertebrate-dominated environment have probably been responsible for the vertebrate-type eye of cephalopods (p. 293).
12. The conclusion that the common adaptive zone shared by cephalopods and fish has been achieved by mutual interactions between the two groups evokes no special evolutionary principles. It assumes that all cephalopod species have (at some period of their evolutionary history) been in competition for food with some vertebrate species and that vertebrates are a source of selection pressures – largely operating through visual behaviour – that maintain and promote convergence upon the ‘fish’ modal type.
13. Selection pressures also operate within behaviour space to maintain and promote the special differences that separate any competitively successful species from all others. Coleoids as a group appear to have retained adaptations associated with such molluscan features as high growth rate and rapid turnover of the population. They are still characteristically crepuscular in habits and have extensive vertical mobility.”
References
O’Dor RK and Webber DM 1986. The constraints on cephalopods: Why squid aren’t fish. Can J Zool 64:1591–1605.
Packard A 1966. Operational convergence between cephalopods and fish: An exercise in functional anatomy. Arch Zool 51:523–542.
Packard A 1972. Cephalopods and fish: The limits of convergence. Biol Rev 47:241–307.
Shigeno S, Sasaki T, Moritaki T, Kasugai T, Vecchione M, Agata K 2008. Evolution of the cephalopod head complex by assembly of multiple molluscan body parts: Evidence from Nautilus embryonic development. J Morphol 269(1):1-17. doi: 10.1002/jmor.10564. PMID: 17654542.
Shigeno S, Sasaki T and von Boletzky S 2010. The origins of cephalopod body plans: A geometrical and developmental basis for the evolution of vertebrate-like organ systems. Pp. 23-34 in Tanabe K., Shigeta Y., Sasaki T and Hirano H (eds) 2010. Cephalopods – Present and Past Tokai University Press, Tokyo.
