
Earlier we looked at
the evolution of the wing in pterosaurs and in bats. Today we’ll look at the evolution of wings in birds. Other than falsifying/modifying the ‘phase shift’ hypothesis (Wagner and Gauthier 1999), there’s nothing heretical about what you’re going to see and read here. Everyone agrees on the taxon list, phylogenic order and bone identification.

Figure 1. The ancestry of birds illustrated by Haplocheirus, Velociraptor, Aurornis, Archaeopteryx and Gallus to scale. Click to enlarge. Thanks to Scott Hartman for his Velociraptor, manus flesh outline oddly omitted.
The origin of feathers and wings
in birds has been well documented in hundreds of publications. Here (Figs. 1, 2) all those accounts have been simplified into just two graphics and a little text.

Figure 2. A selection of pre-bird and bird hands/wings including Haplocheirus, Limusaurus, Velociraptor, Archaeopteryx, Anser, Passer and two versions of the Hoatzin , Opisthocomus, adult and juvenile. Click to enlarge. Not to scale. Note the medial digit of the outlier, Limusaurus, which is a product of neotony, retained from embryonic tissue recapitulating the seven-finger manus of basal tetrapods (figure 3). Note the return of digit 0 fused to the anterior rim of Anser, Passer and the adult Opisthocomus.
Haplocheirus
had grasping hands and trenchant unguals. The fingers were relatively short. Digit 1 was the most robust. Unlike the more basal theropods, Tawa and Herrerasaurus (Fig. 4, 5), manual digits 4 and 5 were absent in Haplocheirus and kin. Digit 3 was also reduced compared to those basal theropods.
Limusaurus
is very much an outlier, not a transitional taxon, different than other related taxa due to its vestigial size and embryonic development. As noted earlier, the Limusaurus manus retains a vestigial embryonic bud of digit ‘0’ which appears in basalmost tetrapods and many embryos, but not otherwise — unless you accept the hypothesis that the anterior process of metacarpal 1 in many extant birds (Fig. 2) is the return of this digit.
Velociraptor
was smaller overall and had longer fingers and longer metacarpals. Note metacarpal 3 is now subequal to metacarpal 2, but metacarpal 1 remains the most robust. One gets the impression that the fingers in Velociraptor had to be stiffer when they supported feathers. At some point they lost or were losing their ability to flex. At the same time the wrist better able to fold the manus in the plane of the forearm, as birds do.
Aurornis
was smaller overall and also had longer more gracile fingers. There is no bow to metacarpal 3 in Aurornis. This manus can be called a wing here.
Archaeopteryx
was overall smaller, but otherwise quite similar to Aurornis. This manus/wing of Archaeopteryx bore large primary feathers.
Anser
is an extant goose. Metacarpal 1 develops an anterior process where digit ‘0’ appeared on Limusaurus. Metacarpal 3 was bowed. The unguals are much smaller. The proximal metacarpals are fused.
Passer
is an extant sparrow. The phalanges are fused to one another.
Opisthocomus
is the extant hoatzin, which goes through a metamorphosis during growth. Juveniles have claws and adults absorb those while fusing the fingers together.
Wagner and Gauthier (1999)
noted the primitive phalangeal formula for tetrapods goes back to Tulerpeton, (Fig. 3) which they considered, “a synapomorphy that arose in the late Devonian, before the origin of Tetrapoda.” Now paleontologists consider Tulerpeton a tetrapod. The phalangeal formula, of course, has roots in Acanthostega, which has three extra digits, one medially and two laterally. Note: it is the reappearance of the medial digit, digit ‘0’, that is key to the present controversy. Note that Tulerpeton has lost one medial and one lateral digit.

Figure 3. Manus of a bird embryo, and two basal tetrapods, Acanthostega and Tulerpeton, the latter with digits 1-3 colorized like the birds in figure 2. Note the extra medial digit in Acanthostega. This is key to the present controversy. The metapterygial axis runs through the longset finger in basal tetrapods.
Embryology
Wagner and Gauthier (1999) report, “There has long been a dissenting view from the hypothesis that the bird hand is composed of digits DI, DII, and DIII. This position is held chiefly by embryologists who argue that the remaining fingers actually represent DII, DIII, and DIV because the DI and DV were thought to have been lost. Morse (19) observed that, when digital reduction occurs in mammals and lizards, the first digit (DI) is invariably the first to be lost in ontogeny, followed by the fifth (DV), and that a modified version of this pattern applies to the foot of birds as well. Thus, the proposition that ultimately became known as Morse’s Law holds that the three functional fingers remaining in adult birds must be DII, DIII, and DIV.”
That hypothesis assumes that the metapterygial axis continued to produce digit 4. The other option is this: Evidently there WAS a phase shift, shifting the metapterygial axis from 4 in basal archosaurs to 3 in basal theropods and birds. This is a possibility that was not considered in prior studies. And it makes sense because theropods lose manual digits 3 and 4.
Sometimes paleontology does not occur out in the field,
or in the lab, but between the ears, as a new way of thinking becomes the solution to a vexing problem. (Note: no DGS was involved in this heretical appraisal.)

Figure 3. The source of the phase shift hypothesis, assuming the homology of manual digit 4 as the first digit to appear on the manus of Alligator (above) and Struthio (the Ostrich, below). Clic to enlarge. It is easy to see how the mistake was made. Evidently there WAS a phase shift, shifting the metapterygial axis from 4 in basal archosaurs to 3 in basal theropods and birds. This is a possibility that was not considered in prior studies.
Which manual digit is the longest in in basal theropods?
Distinct from most other theropods, manual digit 3 is the longest in Herrerasaurus (Fig. 4) and Tawa (Fig. 5). So, digit 3 is where the new metapterygial axis is located on theropods and birds. Digits 4 and 5 are tiny and tinier vestiges, completely lost in later theropods and birds. It doesn’t make sense that the metapterygial axis should produce a vestige – or no digit at all! Rather, it is the metapterygial axis that has shifted one digit medially. That’s the new heretical phase shift promoted here.
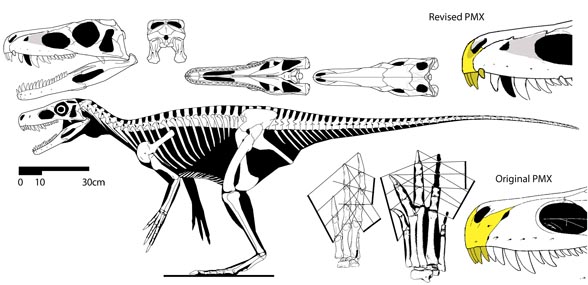
Figure 4. Herrerasaurus. The manus has three functional fingers. The two lateral fingers are vestiges.

Figure 5. The basal theropod, Tawa, with its long manual digit 3. This is where the metapterygial axis has shifted.
Wagner and Gauthier (1999)
also point to the example of the Kiwi manus, some of which have only one finger and two metacarpals. One of these examples had one less phalanx than the other. IMHO you should use fully functioning examples, real wings and real hands, not tiny useless vestiges that are taking various fast tracks toward reduction and disappearance. Wagner and Gauthier also placed the phase shift between Torvosaurus and Allosaurus on their cladogram. That’s an odd place to put a major transition: between two giants. I put the new phase shift at the very base of the Dinosauria, just prior to Herrerasaurus and the basal phytodinosaur, Eoraptor, which also has vestigial lateral fingers.
Wagner and Gauthier (1999) also report,
“We are not aware of any other case in which such a conflict between a developmental and a functional constraint in digit reduction existed.” That’s true. And there is no such conflict in birds if one accepts the novel hypothesis that the metapterygial axis shifted medially as the lateral digits became useless vestiges.
The deeper you get into evolution, the more it all comes together…
References
Müller GB and Alberch P 1990. Journal of Morphology 203, 151–164.
Wagner GP and Gauthier JA 1999. 1,2,3 = 2,3,4: A solution to the problem of the homology of the digits in the avian hand. Proceedings of the National Academy of Science 96:5111-5116.
