
The migration of the central carpals
(the medial and lateral centralia) is today’s topic. We’re going to wonder if these carpals migrated to the medial wrist twice, in mammals and fenestrasaurs (including pterosaurs, Peters 2009).
In basal reptile, like Haptodus (Fig. 1), the two centralia are entirely within the wrist and articulate between the distal and proximal carpals. So that forms our evolutionary baseline.

Figure 1. Carpal evolution from the basal (plesiomorphic) condition represented by Haptodus, to the transitional taxon, Biarmousuchus, to the human carpus where all the bones are renamed (see text). Also note the reduction and disappearance of manual 3.2, m4.2 and m4.3. The two centralia are in gray, absent in humans.
If you’re like me,
the carpal bones are the last ones you learn. They’re small. They’re round. They used to be not that interesting. However, while I’m writing and illustrating this, I’m learning my carpals in a primary fashion and am finding them fascinating.
As you already know…
the carpus evolves in distinct ways in various lineages.
In birds,
a derived arced carpus permits wing folding.
In crocodylomorphs,
the proximal carpals (radiale and ulnare) become greatly elongated.
In tritosaur lizards
The carpus is unossified in Huehuecuetzpalli, likely as the two centralia (in pterosaurs renamed “pteroid” and “preaxial carpal”) were migrating to the medial surface. More on this below.
In legless taxa
While you might think the wrist bones would be among the first to disappear in taxa that have vestigial or absent limbs, that is not so in Adriosaurus, a sister to the ancestor of many snakes.
The synapsid/mammal carpus
In most therapsids (Fig. 1) distal tarsals 4 and 5 become fused. Exceptions include Titanophoneus and Galechirus. The pisiform (in red) becomes enlarged in certain therapsids and mammals (Fig. 4), but is reduced in humans. The medial centralia either disappears (and the prepollex appears de novo) or the medial centralia becomes the prepollex (aka radial sesamoid). The lateral centralia disappears in humans, but not in other primates.
In frogs
The mammal (mole, lemur, elephant, panda) prepollex is not homologous with the amphibian prepollex found in certain frogs. More on that here.
In mammals the carpals change names.
Perhaps this is one other reason why they are the last bones to be learned. Coloring them really helps. Here are the translations.
Proximal Tarsals:
- Radiale = Scaphoid (colored green in all figures here)
- Intermedium = Lunate (amber)
- Ulnare = Triquetrum (pink)
- Pisiform = Pisiform (red)
Centralia
- Medial Centralia = Prepollex (colored gray)
- Lateral Centralia = Lateral Centralia (colored gray)
The question is: is the medial centralia the same bone as the prepollex? The medial centralia of Asioryctes (Fig. 2, a basal mammal) appears to be split in two (which may not be the case in reality), but it retains the configuration seen in Haptodus and Biaromosuchus (Fig.1)
Distal Tarsals:
- DT1 = Trapezium (all colored lavender here)
- DT2 = Trapezoid
- DT3 = Capate
- DT4+5 = Hamate

Figure 2. Asioryctes (basal placental mammal) carpus. Prepollex in gray between scaphoid (=radiale) and trapezium (distal tarsal 1). The ventral scaphoid (= radiale in green) has a ventral tuberosity (upper left). Red oval is the presumed, but missing, pisiform. From Kielen-Jaworowska 1977.
Basal Primate
The hand of Notharctus (Fig. 3), an extinct lemuroid, is transitional between that of Asioryctes and Homo (with many other taxa in-between both!). Note the presence of the prepollex (in gray) and the absence of a medial centralia (like Clark Kent and Superman, you never see them in the same room together). Here the prepollex (medial centralia) is separated from the lateral centralia by a distal tarsal 1 process.

Figure 3. Notharctus (basal primate) wrist elements. The prepollex extends medially (in gray). Otherwise only one centralia appears here (also in gray), if the prepollex is not the medial centralia.
Pre-panda and panda thumbs
Simocyon (Salesa et al. 2005, a Miocene pre-panda) has a prepollex (radial sesamoid) and Ailuropoda (giant panda, Fig. 4) expands this to create a “false thumb” extending outside of the wrist proper. The similarity in morphology to the carpus of Notharctus (Fig. 3) is notable. Apparently in pre-pandas the lateral centralia became lost or fused to the radiale (= scaphoid, compare to Figure 3). However, the medial centralia (prepollex, radial sesamoid, “rs,” in gray) is radically enlarged in the red panda (Ailuropoda). This may not be a sesamoid, but a centralia after migration, as in pterosaurs.
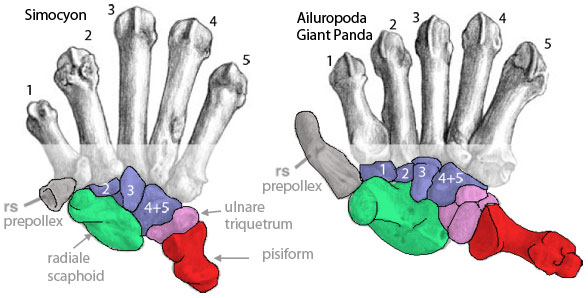
Figure 4. Metacarpals and carpals of Simocyon (Miocene) and Ailuropoda (Recent) from Salesa et al. 2005. Note the enlargement of the prepollex (radial sesamoid, gray) and otherwise the lack of centralia. The enlargement of the pisiform is interesting and potentially confusing, but not pertinent to the present discussion as it emerges from the lateral wrist. Not sure what this means, but Simocyon does not have a distal tarsal 1.
Comparisons to fenestrasaurs (including pterosaurs)
Earlier we discussed how the medial centralia becomes the pterosaur pteroid and the lateral centralia becomes the preaxial carpal after these migrate to the medial wrist rim from their central origins (Fig. 5, Peters 2009). This is heretical thinking, of course, but now this migration hypothesis has support in that it may be convergent with the appearance of the prepollex in Notharctus and other mammals.

Figure 5. Click to enlarge. The origin of the pterosaur wing. Note the disappearance of the centralia and the reappearance of the pteroid and preaxial carpal.
I learned something while writing this.
Let me know your thoughts, especially if I missed any interesting configurations or evolutions.
References
Hamrick MW and Alexander JP 1996. The Hand Skeleton of Notharctus tenebrous (Primates, Notharctidae) and Its Significance for the Origin of the Primate Hand. American Museum Novitates 3182, 20pp.
Kielan-Jaworowska Z 1977. Evolution of the therian mammals in the Late Cretaceous of Asia. Part n. Postcranial skeleton in Kennalestes and Asioryctes. In: Z. Kielan-Jaworowska (ed.) Results Polish Mongolian Palaeont. Expeds. VIII. – Palaeont, Polonica, 37, 65-84.
Peters D 2009. A reinterpretation of pteroid articulation in pterosaurs. Journal of Vertebrate Paleontology 29:1327-1330.
Salesa MJ, Antón M, Peigné S and Morales J 2005. Evidence of a false thumb in a fossil carnivore clarifies the evolution of pandas. Proceedings of the National Academy of Sciences of the United States of America. abstract and pdf
