
Lacalli 2024 looked at the origin of chordate somites
(= torso muscles also found in fish that enable wriggling for propulsion) by reexamining Middle Cambrian Pikaia (Figs 1–3).
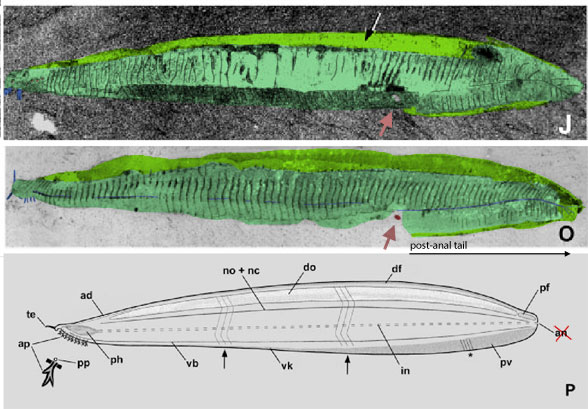
Figure 1. Above: Pikaia in situ. Colors added here. Below: Original diagram.
Lacalli 2024 misunderstood Pikaia
because he did not have a relevant Bauplan (Figs 2) to compare it to. Examples follow.
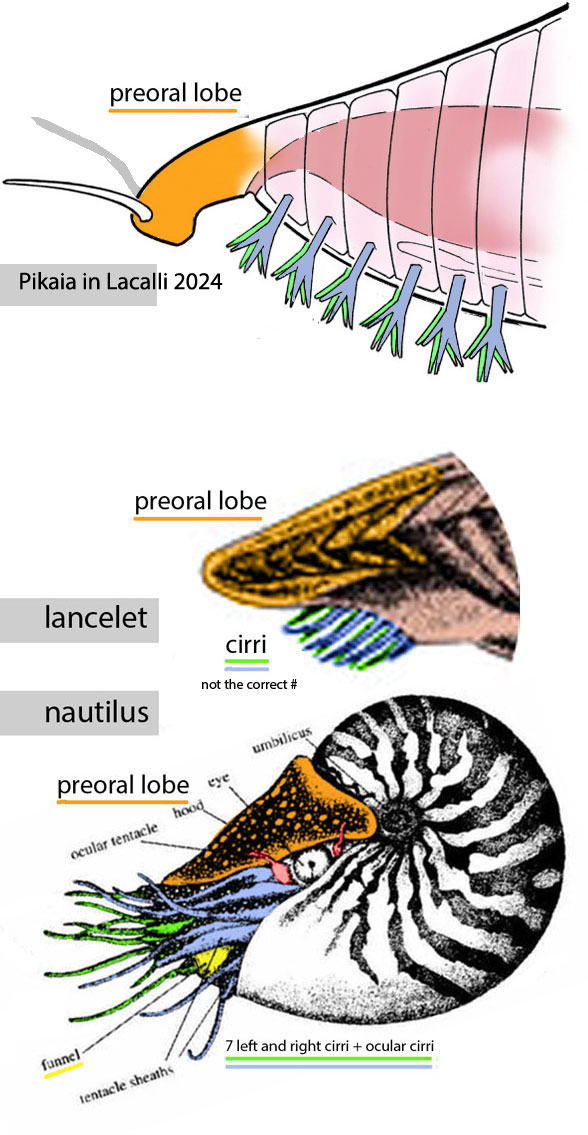
Figure 2. Pikaia compared to a lancelet and a nautilus.
Caption in Lacalli 2024 describing figure 1.
“The anterior end of Pikaia, oriented with the mouth facing down. The anterior appendages (ap) are generally interpreted as gills or gill-related structures, and repeat in register with the anterior segments (pink).
Those are not “gills or gill-related structures.” Those appendages are buccal cirri as in related lancelets and nautili. They are feeding structures. Gills are internal.

Figure x. Lancelet buccal cirri, evolved from the time of Pikaia.
“The pharynx, dorsal organ and ventral blood vessel are shown in outline. The point of the diagram is to show that there is no evidence for appendages positioned forward of the putative somite series, so if the latter indicates the beginning of the trunk, the region constituting what is effectively the head of Pikaia is exceedingly small.”
The preoral hood present in Pikaia and related taxa is overlooked by the author
Lacalli 2024 concluded,
“If Pikaia is a chordate, then even without a notochord it would represent a comparatively advanced member of that lineage based on the way its body is organized and the mode of swimming that organization implies.”
Phylogenetic analysis recovers last common ancestors preceding every node. That alone would determine if Pikaia was a chorate, even if it did not have a notochord, though that would be well neigh impossile.
“Pikaia is nevertheless difficult to place in relation to living chordates.
By contrast, Pikaia is easy to place in the LRT.
“It has, on the one hand, structures with no obvious chordate counterpart (e.g., the dorsal organ and the apparent head shield, or “anterior unit”, while on the other, the evidence for structures it should have, like the notochord and nerve cord, is equivocal.”
This is not true. One can demonstrate (Figs 1–3) all the elements are homologous with structures in lancelets and related taxa.
“There are nevertheless questions concerning chordate origins that one can address using Pikaia as a point of reference, with that of somite origins being the most obvious. The problem, in essence, is how to convert a large coelom spanning the trunk to a repeating series of smaller coelomic compartments.”
Segmentation occurs in invertebrates and vertebrates.

Figure 3. Extant lancelet (genus: Amphioxus) in cross section and lateral view. The gill basket nearly fills an atrium, which intakes water + food, sends the food into the intestine and expels the rest of the water.
“Ruppert’s ideas on the hemichordate collar cord provide a useful starting point by highlighting the role of structures of this type in innervating muscles distant from the body surface.”
The collar cord is not useful. It is a derived trait.
“The extension of an initial, anterior invagination, to form an extended neural tube, would follow as the targets of innervation, the myotomes, were themselves replicated as an extended anteroposterior series.
Marine nematodes, like Enoplus meridionalis, provide good ancestral taxa.
“In this scenario, the neural tube can be seen as an innovation at the anatomical level that solves a histological problem, of the inherent limitations of intraepithelial innervation”
That’s how the embryo develops, even in humans.
“Any chordate with somites would then be expected to have a neural invagination comparable to the collar cord that extends as far along the anteroposterior axis as the somites themselves, or a neural tube of comparable length.”
Wrong Bauplan.
“The argument for homology between the collar cord and the chordate nerve cord remains open, but the case in favor is supported by evidence at the cellular and molecular level of how similar the nervous systems of hemichordates and protochordates are, with vertebrates being, in some respects, an outlier.”
The author has it backwards. Hemichordates are derived, not primitive.
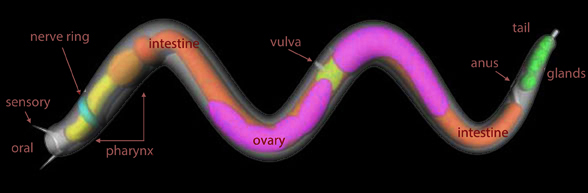
Figure 4. Illustration of an enoplid nematode with labels.
“The position of the neural tube in ancestral chordates would have depended on whether a notochord was present or not, which then begs the question of how the body was supported in the absence of a notochord.”
Add taxa. Looks at Enoplus, the nematode (Fig 4). Don’t speculate. Observe.
“Since improvements to the locomotory abilities and efficiency of ancestral chordates can be assumed to have been progressive, it is by no means clear that a structural support as substantial as a notochord would have been required in the first instance to act as a compression strut.”
Worms can’t swim. Lancelets can swim. That’s what a notochord gives to a taxon.
“Whether Pikaia has a notochord or supports its body by other means remains an unresolved problem. If the dorsal organ, whatever its tissue of origin, is in some way concerned with body support, then Pikaia is potentially representative of a transitional step in the sequence by which chordates evolved body support systems of increasing efficiency.”
Leaving the sea floor is what turns a nematode into a lancelet.
“And, if the dorsal organ is not simply an overlarge notochord, one would have to conclude that a notochord of typical chordate type evolved considerably later than the somite series.”
Probably in lock-step with one another.
“This would place the dorsal organ at center stage as an early but temporary solution to the problem of body support.”
Make that a permanent solution in a structure similar to or identical to cartilage. So body support once wriggled left, snaps back right in a continuous sine wave of open water locomotion.
“An alternative, that Pikaia represents an independent, now extinct chordate lineage with somites but no notochord, is also possible, or it could once have had a notochord that has been lost as its functions were taken over by the later evolution of the dorsal organ. There remain, in sum, a number of unresolved questions concerning this remarkable fossil and how it is to be interpreted.”
The terms ‘preoral hood; ‘cirri’, ‘lancelet’, ‘myxiia’, ‘lamprey, ‘hagfish’ and ‘nematode’ are not mentioned in the text,
The term ‘amphioxus’ is mentioned several times.
“With regard to amphioxus specifically, there is a lingering misconception that the burrowing habit has resulted in its becoming a rather poor and ineffective swimmer. The contrary is, in fact, the case, as anyone who has tried to net an amphioxus swimming at peak speed will attest. The animal is mass of muscle and connective tissue capable of powerful writhing movements when restrained, and of considerable speed when swimming, greater than many fish when adjusted for relative body size.”
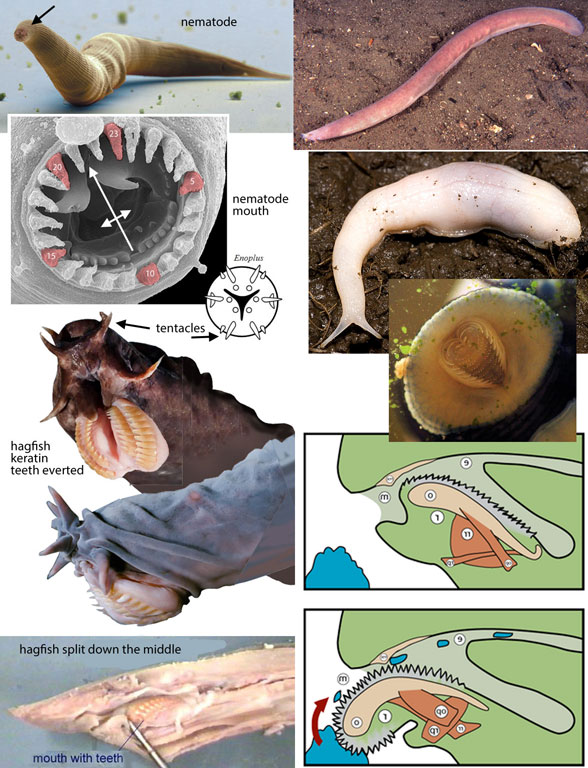
Figure 5. Nematodes, hagfish and slugs have so many traits in common, one wonders why they are not related to one another.
Localli 2024 followed the outdated enteropneust Bauplan model.
Enteropneusts are derived lancelets with fewer, weirder parts. Better to use taxa related to Pikaia in the LRT. These are recovered by adding taxa.
References
Bardack D and Richardson ES Jr 1977. New aganathous fishes from the Pennsylvanian of Illinois. Fieldiana Geology 33(26):489–510.
Linnaeus C 1758. Systema naturæ per regna tria naturæ, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Tomus I. Editio decima, reformata.
Lacalli T 2024. The Cambrian fossil Pikaia, and the origin of chordate somites. BMC EvoDEvo 15:1.
Mallatt J and Holland ND 2013. Pikaia gracilens Walcott: stem chordate or already specialized in the Cambrian? Journal of Experimental Zoology, Part B, Molecular and Developmental Evolution 320B: 247–271.
Walcott CD 1911. Cambrian geology and paleontology. Middle Cambrian annelids. Smithsonian Miscellaneous Collections 57(5):109–144.
Prior posts on lancelets and relatives:

Dear Mr. Peters,
You should be aware that the siphuncle in cephalopod molluscs (such as the Nautilus) is not an analogous structure to the notochord as seen in chordates (such as the Lancelet), as outside their general shape, they are entirely different anatomically and in terms of function. A siphuncle is an internally hollow organ responsible for controlling buoyancy, and an important part of the circulatory system, composed of soft tissue and blood vessels. In the meantime, a notochord, even in its most primitive extant form, is composed of a protein core, encased with a pair of helical sheaths, composed of collagen fibres, forming an attachment to the muscular system.
Best regards.
I see no citation to your claims, Mr/Ms Anonymous. So let’s help the readers…
According to Greenwald, Cook and Ward 1982, “The siphuncle of the chambered nautilus (Nautilus macromphalus) is composed of a layer of columnar epithelial cells resting on a vascularized connective tissue base. The siphuncular epithelium taken from chambers that have not yet begun to be emptied of cameral liquid has a dense apical brush border. The great number of apical cell junctions (zonula adherens) compared to the number of nuclei suggests extensive interdigitation of these cells. The perinuclear cytoplasm of these preemptying cells is rich in rough endoplasmic reticulum. The siphuncular epithelium of both emptying and “old” siphuncle (which has already completed emptying its chamber) both show little rough endoplasmic reticulum but do contain extensive systems of mitochondria-lined infoldings of the basolateral plasma membranes. Active transport of NaCl into the extracellular space of this tubular system probably entrains the water transport involved in the chamber-emptying process. Both emptying and old siphuncular epithelium also show large basal infoldings (canaliculi) continuous with the hemocoel, which appear to be filled with hemocyanin. The apical cell junctions of emptying and old siphuncular epithelium contain septate desmosomes that may help to prevent back-flow of cameral liquid into the chambers.”
“Persistent notochordal canal” is a pathological condition in humans according to Christopherson et al 1999. Does that occur in more basal chordates? I don’t know, but it is a clue we can follow.
You wrote, “even in its most primitive extant form,”.
This split of chordates and cehalopods, which remains a hypotheses here, is proposed to have occurred in the last common ancestor of hagfish and nautilus. I am not aware of that LCA has an extant form. But that should not stop us…
According to a UChicagoMedicine website from 2019 speaking on Tethymyxine a Cretaceous hagfish fossil, “They don’t have eyes, or jaws or teeth to bite with, but instead use a spiky tongue-like apparatus to rasp flesh off dead fish and whales at the bottom of the ocean.”
If that sounds familiar, that may be because, “The mouth structures of many molluscs include a specially adapted rasp-like tongue called a radula.”
You might have mentioned the eye is not the same either. Nautilus has a pinhole, not a lens as in vertebrates and squid. To your point, you can find this phrase online: “Hagfish eyes also lack a lens and pigment.”
You said, “the siphuncle in cephalopod molluscs (such as the Nautilus) is not an analogous structure to the notochord as seen in chordates.” Did you mean to say ‘homologous structure’? That’s what the presented hypothesis proposes: homologous. Clearly hagfish and nautilus have diverged since those last common ancestor days. Whatever they are now, the presented hypothesis promotes the idea that the siphuncle and notochord started off as the same structure in the last common ancestor of chordates and cephalopods. Presently that last common ancestor is unknown, but the presented clues point in this direction.
Your job, as a scientist, is to find that last common ancestor and show that it does not have a structure that will someday become a notochord in one clade and a siphuncle in the other. It is not enough to say they are different now. They clearly are, just as the notochord in humans is different from that of lancelets.
It’s an idea worth investigating. Let us know of your progress! Don’t omit relevant taxa.
your tree doesn’t include the taxa that the authors here proposed to be the ancestors of Chordates – Yunnanozoon and Vetulicolians
Correct! Largely because they don’t have skeletons. Here’s where those taxa were discussed. Hope this helps.