Figure 1. Australohyaena, a big sister to the cat-like marsupial, Oxyaena, in the LRT. That’s on oddly low jaw glenoid. Distinct from other marsupials, the jugal might NOT have extended to the jaw joint. Originally the frontals were considered nasals.
Australohyaena antiqua(Forasiepi et al. 2014; Late Oligocene; UNPSJB PV 113; Fig. 1) was originally nested with Borhyaena, but here nests with a previously untested taxon, Oxyaena (Fig. 1). Australohyaena is larger and more robust, but the orbit is no larger. The horizontal bar jaw joint makes the jaw a simple hinge with not transverse action possible. The jaw joint falls below the tooth row. The frontals were originally considered nasals.
If you think
marsupial Australohyaena looks like a placental lion (genus: Panthera leo), I agree with you (Fig. 2).
Figure 2. Panthera leo skull and skeleton. This taxon nests basal to hyenas + wolves.
References Forasiepi AM, Babot MJ and Zimicz N 2014.Australohyaena antiqua (Mammalia,
Metatheria, Sparassodonta), a large predator from the Late Oligocene of Patagonia, Journal of Systematic Palaeontology, DOI: 10.1080/14772019.2014.926403
The traditional eutherian clade
‘Taeniodonta‘ is polyphyletic in the large reptile tree (LRT, 1365 taxa, subset Fig. 4). Earlier here and here are parts 1 and 2 of this series.
Figure 1. Conoryctes fossil and drawing from Schoch 1986.
Conoryctes comma(originally Hexadon molestus, Cope 1881; Schoch 1986; Paleocene; AMNH 3395). Here the Schoch drawings were not a great match for the fossil data. This taxon nests with the much earlier, Early Cretaceous marsupial, Vincelestes(Fig. 4). These were derived from Huerfanodon (Fig. 5). Note the elevated premaxila, flattened molars and deep dentary. The Schoch 1986 diagnosis of this genus is based on dental traits. He also compared Conoryctes to Huerfanodon, a related taxon in the LRT, but he also compared them to Onychonycteris, an unrelated, but convergent eutherian close to phenacodontids (Fig. 3), which is close to Conoryctella (Fig. 6) another putative taeniodont.
Figure 3. Subset of the LRT labeling several traditional taeniodonts in red, indicating the traditional clade Taeniodonta is polyphyletic and should therefore be abandoned.
Conoryctes is so unpopular,
it has no Wikipedia entry.
Figure 4. Vincelestes soul showing the separation of the nasals and frontals by the conjoined maxillae housing giant canine roots, as in sister Thylacosmilus.
Vincelestes neuquenianus(Bonaparte 1986, Early Cretaceous, 130 mya). Derived from a sister to Huerfanodon, Vincelestes is a carnivorous marsupial sister to the traditional taeniodont, Conoryctes. Note the hyper-enlarged canines and short rostrum. Premolars were not carnassial in shape, but still able to process by cutting and grinding. Nine individuals are known. Skeleton is probably a chimaera of several specimens and the degree of completion is unknown. The tail is extraordinarily long and provided with deep chevrons. Image from Digimorph.org and used with permission. Scale bar = 1 cm.
Figure 5. Heurfanodon skull. This late survivor of a Jurassic radiation is from the Eocene. It is transitional between didelphids and the Vincelestes-Thylacosmilus clade.
Huerfanodon torrejonius(Cope 1882; Eocene; AMNH 3224, Schoch 1986, USNM 15412) was traditionally considered a taeniodont, but here nests basal to the Vincelestes/ Thylacosmilus clade. Despite its late appearance, this taxon is more primitive than the others, which puts its genesis deep into the Jurassic. In dorsal view the skull is not compressed anterior to the jugals, similar to the ancestral Chironectes.
Figure 6. Conoryctella pattersoni nests with the Eutherian herbivore Onychonycteris in the LRT, not with the marsupial Conoryctes, according to the LRT, based on the present data.
References Cope ED 1881. On some Mammalia of the lowest Eocene beds of New Mexico. Palaeontological Bulletin 33:484-495. Schoch RM 1986. Systematics, functional morphology and macroevolution of the extinct mammalian order Taeniodonta. Bulletin of the Peabody Museum of Natural History, Yale University, New Haven. 307pp.
You can find part 1
of “‘Taeniodonta’ is polyphyletic” here. Former ‘taeniodonts’ nest is a variety of nodes in the LRT (see Fig. 5), not as a monophyletic clade.
Figure 1. Showalteria. Not much there. Adding more rounds out the skull, a likely marsupial relative of the sabertooth marsupial, Thylacosmilus in figure 2.
Schowalteria clemensi (Fox and Naylor 2003; Maastrichtian, Cretaceous, 68mya) was considered a stylinodont, taeniodont eutherian, but here nests with the marsupial sabertooth, Thylacosmilus (Fig. 2), if the bones above have been correctly identified. Otherwise, Thylacosmilus has no close relatives in the LRT. A traditional close relative, Patagosmilus, was shifted away earlier.
Figure 2. Thylacosmilus skull. Note the deep maxillae in dorsal contact containing giant canine roots. These are not present in Patagosmilus.
Thylacosmilus atrox (Riggs 1933; Miocene, 40-3 mya; 1 m long) was a leopard-like predator, but plantigrade derived from a sister to Schowalteria. The premaxillary teeth are absent here, perhaps lost during taphonomy. The canines are enlarged to curved fangs. Their roots extend above the orbits. The dentary has ventral processes that guide and protect them. The lower canines are round stubs. Tiny lower incisors appear between them. The coronoid process is small, indicating a wide, but weak bite. Both Thylacosmilus and Schowalteria were derived from a sister to Vincelestes (Fig. 3) in the Early Cretaceous.
Figure 3. Vincelestes soul showing the separation of the nasals and frontals by the conjoined maxillae housing giant canine roots, as in sister Thylacosmilus.
Vincelestes neuquenianus (Bonaparte 1986, Early Cretaceous, 130 mya). Derived from a sister to Huerfanodon(Fig. 5), Vincelestes is a carnivorous marsupial sister to the traditional taeniodont, Conoryctes. Note the hyper-enlarged canines and short rostrum. Premolars were not carnassial in shape, but still able to process by cutting and grinding. Nine individuals are known. Skeleton is probably a chimaera of several specimens and the degree of completion is unknown. The tail is extraordinarily long and provided with deep chevrons. Image from Digimorph.org and used with permission. Scale bar = 1 cm.
Figure 4. Heurfanodon skull. This late survivor of a Jurassic radiation is from the Eocene. It is transitional between didelphids and the Vincelestes-Thylacosmilus clade.
Huerfanodon torrejonius(Cope 1882; Eocene; AMNH 3224, Schoch 1986, USNM 15412) was traditionally considered a taeniodont, but here nests basal to the Vincelestes/ Thylacosmilus clade. Despite its late appearance, this taxon is more primitive than the others, which puts its genesis deep into the Jurassic. In dorsal view the skull is not compressed anterior to the jugals, similar to the ancestral Chironectes.
Figure 5. Subset of the LRT labeling several traditional taeniodonts in red, indicating the traditional clade Taeniodonta is polyphyletic and should therefore be abandoned.
References Bonaparte JF 1986. Sobre Mesungulatum houusayi y nuevos mamíferos Cretácicos de Patagonia, Argentina [On Mesungulatum houssayi and new Cretaceous mammals from Patagonia, Argentina]. Actas del IV Congreso Argentino de Paleontología y Biostratigrafía 2:48-61. Cope ED 1881. On some Mammalia of the lowest Eocene beds of New Mexico. Palaeontological Bulletin 33:484-495. Cope ED 1882. Contributions to the history of the Vertebrata of the lower Eocene of Wyoming and New Mexico, made during 1881. Proceedings of the American Philosophical Society 20(111):139-197. Fox RC and Naylor BG 2003. A Late Cretaceous taeniodont (Eutheria, Mammalia) from Alberta, Canada. Neues Jahrbuch für Geologie und Paläontologie. 229 (3):393–420. Riggs EC 1934. A new marsupial saber-tooth from the Pliocene of Argentina and its relationships to other South American predacious marsupials. Transactions of the American Philosophical Society 24, 1–32. Schoch RM 1986. Systematics, functional morphology and macroevolution of the extinct mammalian order Taeniodonta. Bulletin of the Peabody Museum of Natural History, Yale University, New Haven. 307pp.
Rule #1: More taxa more precisely nest all taxa Once again, latest Cretaceous Cimolestes goes under review, this time with many more candidate sister taxa. At present the best material for this genus is a single mandible with a complete set of teeth (Fig. 1). Rook and Hunter 2013 nest Cimolestes as the direct outgroup to the tradtional clade Taeniodonta.
According to Wikipedia,
“[Members of the genus Cimolestes] were once considered to be marsupials, then primitive placental mammals, but now are considered to be members of the order Cimolesta (which was named after the genus), outside of placental mammals proper (Rook and Hunter 2013). Before they were determined to be non-placental eutherians, the cimolestids were once considered the common ancestral group of the clades Carnivora and the extinct Creodonta.”
Figure 1. Cimolestes is represented by a toothy mandible. Here it nests with the extant Dasyurus if the back of the skull is shorter. Apparently the coronoid process is oddly narrow. I have not seen incisors like this in any other mammal, but Dasyurus comes close.
With so few traits to score, Cimolestes is difficult to nest and generally causes loss of resolution, especially when other taxa data include skulls without mandibles (so, no comparable traits = loss of resolution). When deleting taxa without preservation of the mandibles the best match is with the extant marsupial, Dasyurus (Fig. 1), but with a narrower coronoid process and larger incisors, and therefore a likely shorter and smaller cranial region.
Figure 2. Traditional Taeniodonta in a cladogram from Rook and Hunter 2013. Colors and list of body parts added here. With more taxa to be attracted to (1362 in the LRT) this clade splits up according to the three colors shown here.
Traditionally Cimolestes is considered a basal taeniodont and all taeniodonts are considered eutherians (placentals). Other traditional taeniodonts include Protictis, Onychodectes and Stylinodon. The Rook and Hunter cladogram of eutherian relationships nests only one traditional taeniodont alongside Cimolestes (Fig. 3) and the basalmost member of the tenrec-odontocete clade (in the LRT), Anagale.
Figure 3. The Rook and Hunter cladogram that nested traditional Taeniodonts within their Eutheria. Colors and tones added here for clarity and comparison. The LRT does not confirm most of these relationships.
In the large reptile tree (LRT, 1362 taxa, subset Fig. 4) none of these taxa nest with one another. Their previous joining may be due to eyeballing, a reliance on dental traits and taxon exclusion. That’s all Cope had available at the time. Modern workers appear to have followed traditional taxon lists and convergent dental traits without testing a wider gamut of taxa. The LRT includes more taxa and does not emphasize dental traits.
When tested with additional taxa,
(Fig. 4) the traditional eutherian clade Taeniodonta is polyphyletic and should be abandoned. Only a few traditional members are closely related to one another.
Figure 3. Subset of the LRT labeling several traditional taeniodonts in red, indicating the traditional clade Taeniodonta is polyphyletic and should therefore be abandoned.
As typical, taxa in the LRT provide and document a gradual accumulation of derived traits that competing cladograms cannot match.
More former taeniodonts to come.
References Fox RC 2015. A revision of the Late Cretaceous–Paleocene eutherian mammal Cimolestes Marsh, 1889. Canadian Journal of Earth Sciences (advance online publication) doi: 10.1139/cjes-2015-0113. Marsh OC 1889. Marsupialia, Cimolestidae. American Journal of Science and Arts 3d ser., XXXVIII, 89, pl. iv, figs. 8–19. Rook DL and Hunter JP 2013. Rooting around the eutherian family tree: the origin and relations of the Taeniodonta. Journal of Mammal Evolution
DOI 10.1007/s10914-013-9230-9
Traditionally sabertooth marsupials nest together with other carnivorous marsupials in a clade Cope 1875 called Creodonta and Ameghino 1895 called Saprassodonta (back when creodonts were considered archaic placentals).
Figure 1. Patagosmilus to scale alongside Hadrocodium. These sabetooth taxa are not directly related to Thylacosmilus in the LRT. Note the shallow rooted canine. Note the first molar is now the last premolar, contra the original drawing interpretation. The premaxilla is hypothetical based on phylogenetic bracketing and not scored. At a screen resolution of 72 dpi (standard) these are full scale images.
Patagosmilus goini(Fig. 1; Forasiepi and Carlini 2010) is a large sabertooth marsupial from the Middle Miocene of south America traditionally considered a sister to the more famous and distinctly different and more famous sabertooth from South America, Thylacosmilus (Fig. 2). After testing in the large reptile tree (LRT, 1361 taxa), Patagosmilus nests with the ultra tiny basal sabertooth metatherian, Hadrocodium (Figs. 1,3) from the Early Jurassic. Evidently transitional taxa have yet to be discovered (or tested).
Figure 2. Thylacosmilus skull. Note the deep maxillae in dorsal contact containing giant canine roots. These are not present in Patagosmilus.
A little backstory: Thylacosmilus atrox(Riggs 1933; Miocene, 40-3 mya; 1 m long) was a leopard-like predator, but plantigrade. Thylacosmilus was a sister to Early Cretaceous Vincelestes. The premaxillary teeth are either absent or were taphonomivally lost here. The canines are enlarged to curved fangs. Their long roots extend above the orbits. The dentary has deep ventral processes that guide and protect the canines. The lower canines are round stubs. The coronoid process is small. The mandible was able to open nearly 90º.
Figure 3. Tiny Hadrocodium (Early Jurassic) nests as a sister to Patagosmilus (middle Miocene) in the LRT.
Hadrocodium wui(Luo, Crompton and Sun 2001; Early Jurassic; skull length: 1.2cm), known only from a tiny skull about the size of a thumbnail, Hadrocodium was originally considered a juvenile basal mammal, but later a tiny adult. Hadrocodium has a relatively larger brain size and more advanced ear structure than Megazostrodon. Hadrocodium nests with other basal metatherians with three (not two) molars, Morganucodon and Volaticotherium. The first molar was originally considered the third premolar.
Figure 4. Vincelestes soul showing the separation of the nasals and frontals by the conjoined maxillae housing giant canine roots, as in sister Thylacosmilus.
Contra traditional studies none of these taxa are related to marsupial creodonts and/or borhyaenids, like Borhyaena and Hyaenodon (Fig. 5), all of which have large canine teeth, none of which have saber teeth.
Figure 5. Subset of the LRT focusing on the Metatheria. Here sabertooth Patagosmilus nests far apart from sabertooth Thylacosmilus, which nests apart from the clade of borhyaenid marsupials. A red square is placed next to Carnivora to indicate the presence of Smilodon, Haplophoneus and other sabertooth cats.
Oddly, elephants, walruses, deer and other taxa with hyper-elongated teeth are never considered sabertooth proboscidians, sabertooth seals and sabertooth deer. Perhaps this is so because such teeth have a round cross-section, not a narrow, sword/saber-like morphology.
References Ameghino F 1892. (Issued in 1894.) Enumeration synoptique deses pesces de mammiferes fossilesdes formation sócénes de Patagonie. Boletindela, Academia Nacionalde Cienciasen Cordoba, XIII, p.259 (p.108 in reprint). Cope ED 1875. On the Supposed Carnivora of the Eocene of the Rocky Mountains. Proceedings of the Academy of Natural Sciences, Philadelphia. pp. 444–449. Forasiepi AM and Carlini AA 2010. A new thylacosmilid (Mammalia, Metatheria, Sparassodonta) from the Miocene of Patagonia. Zootaxa. 2552, ss. 55–68.
Keep in mind the concept of convergence whenever reviewing purported members of the Creodonta. Several purported creodonts have been added to the large reptile tree (LRT, 1342 taxa) recently.
According to Wikipedia, “Creodonts were the dominant carnivorous mammals from 55 to 35 million years ago, peaking in diversity and prevalence during the Eocene.”
McKenna1975 considered the Creodonta
the sister taxa to the Carnivora within the clade Ferae (Carnivora + Pholidota (= pangolins)). The LRT finds pretty much the same relationship, but with creodonts on the marsupial side of the node and carnivores on the placental side of the node. The arboreal didelphid Caluromys is the only taxon that nests between marsupial creodonts and placental carnivores at present. (In the LRT pangolins nest with currently dissimilar bats. Ancestors of both, Chriacus and Zhangheotheriumwere much more similar.)
Halliday et al. 2015 nested creodonts
as sisters to pangolins in a cladogram that bore little to no resemblance to the LRT.
Figure 1. Oxyaena, a traditional creodont. This is a cat-like member of the carnivorous Marsupialia.
According to Wikipedia “Creodonta was coined by Edward Drinker Cope in 1875. Cope included the oxyaenids and the viverravid Didymictis but omitted the hyaenodontids. In 1880. he expanded the term to include Miacidae, Arctocyonidae, Leptictidae (now Pseudorhyncocyonidae), Oxyaenidae, Ambloctonidae and Mesonychidae. Cope originally placed creodonts within the Insectivora. In 1884, however, he regarded them as a basal group from which both carnivorans and insectivorans arose.Hyaenodontidae was not included among the creodonts until 1909. Over time, various groups were removed, and by 1969 it contained, as it does today, only the oxyaenids and the hyaenodontids.”
Figure 2. Hyaenodon horridus was the size of a large dog. This carnivorous marsupial is considered a traditional creodont.
The LRT recovers members of the traditional Creodonta in the carnivorous clade of the Marsupialia (Fig. 3). Earlier we looked at similar situation with members of the Didelphidae.
Figure 3. Subset of the LRT focusing on Basal Mammalia including Creodonta. Members of the Didelphidae and Creodonta are sprinkled throughout this subset.
Here (Fig. 3) the traditional creodont Sinopa (Fig. 4) nests with the extant dasyurids, Dasyurus and Sarcophilus (Fig. 4).
Figure 4. Adding Sinopa to the LRT nests it here, between the extant quoll (Dasyurus) and the extant Tasmanian devil (Sarcophilus).
References Andrews CW 1906. Descriptive Catalogue of the Tertiary Vertebrata of the Fayum, British Museum. Cope ED 1880. On the Genera of the Creodonta. Proceedings of the American Philosophical Society. 19(107): 76–82. Halliday TJD, Upchurch P and Goswami A 2017. Resolving the relationships of Paleocene placental mammals. Biological Reviews: n/a–n/a. doi:10.1111/brv.12242. ISSN 1464-7931. Matthew WD 1901. Additional Observations on the Creodonta. Bulletin of the American Museum 14:1. McKenna MC 1975. Toward a phylogenetic classification of the Mammalia. Pp. 21–46 in Luckett WP and Szalay FS. Phylogeny of the Primates. New York: Plenum. Morlo M, Gunnell G and Polly PD 2009. What, if not nothing, is a creodont? Phylogeny and classification of Hyaenodontida and other former creodonts. Journal of Vertebrate Paleontology 29(Supplement 3): 152A. Polly PD 1994. What, if anything, is a creodont?. Journal of Vertebrate Paleontology. 14: 42A. Sinclair WJ 1905. The Marsupial Fauna of the Santa Cruz Beds, Proceedings of the American Philosophical Society 49:73. Wortman JL 1901-1902. Eocene Mammalia in the Peabody Museum, pt. i. Carnivora,” American Journal of Science 11–14.
Smith 2011 reported, at the beginning of the Eocene, 55mya, “the diversity of certain mammal groups exploded.” These “modern mammals”, according to Smith, ‘ consist of rodents, lagomorphs, perissodactyls, artiodactyls, cetaceans, primates, carnivorans and bats. Although these eight groups represent 83% of the extant mammal species diversity, their ancestors are still unknown. A short overview of the knowledge and recent progress on this research is here presented on the basis of Belgian studies and expeditions, especially in India and China.’
Contra the claims of Smith 2011
in the large reptile tree (LRT, 1354 taxa, subsets Figs. 2–4) prototherians are known from the late Triassic (Fig. 1). Both metatherians and eutherians are known from the Middle Jurassic. Many non-mammal cynodonts survived throughout the Mesozoic. In addition, the ancestors of every included taxon are known back to Devonian tetrapods.
Noteworthy facts after an LRT review (Fig. 1):
All known and tested Mesozoic mammals (Fig. 1) are either small arboreal taxa or small burrowing taxa (out of sight of marauding theropods).
All Mesozoic marsupials are more primitive than or include Vintana (Late Cretaceous).
All Mesozoic placentals are more primitive than Onychodectes (Paleocene).
Figure 1. Select basal cynodonts and mammals set chronologically. The divergence times for placentals (Eutheria), marsupials (Metatheria) and monotremes (Mammalia) are estimated here.
Given those parameters
we are able to rethink which mammals were coeval with dinosaurs back on phylogenetic bracketing (= if derived taxa are present, primitive taxa must have been present, too).
Smith reports,“The earliest known mammals are about as old as the earliest dinosaurs and appeared in the fossil record during the late Trias around two hundred and twenty million years ago with genera such asSinoconodon (pre-mammal in the LRT), Morganucodon(basal therian in the LRT) andHadrocodium (basal therian in the LRT). However, the earliest placental mammals (Eutheria) were not known before the Early Cretaceous.Eomaia scansoria (not eutherian in the LRT) from the Barremian of Liaoning Province, China is the oldest definite placental and is dated from a hundred and thirty million years ago.”
Mesozoic Prototherians
All included fossil taxa are Mesozoic. Two others are extant (Fig. 2).
Figure 2. Mesozoic prototherians + Megazostrodon, the last common ancestor of all mammals. Only two taxa (gray) are post-Cretaceous.
Derived Didelphodon is Late Cretaceous, which means sisters to Thylacinus through Borhyaena were also present in the Mesozoic.
Derived Vintana is Late Cretaceous, which means sisters to herbivorous marsupials were also present in the Mesozoic.
Figure 3. Mesozoic metatherians (in black), later taxa in gray. Whenever derived taxa are present in the Mesozoic (up to the Late Cretaceous) then ancestral taxa, or their sisters, were also present in the Mesozoic. Didelphis is extant, but probably unchanged since the Late Jurassic/Early Cretaceous.
Mesozoic Eutherians (= Placentals)
Rarely are placental mammals identified from the Mesozoic, because many are not considered placentals.
Placentals (in the LRT) are remarkably rare in the Mesozoic, but sprinkled throughout the cladogram, such that all taxa more primitive than the most derived Mesozoic taxon (Anagale and derived members of the clade Glires, Fig. 4, at present a number of multituberculates) must have had Mesozoic sisters (Carnivora, Volitantia, basal Glires).
Figure 4. Mesozoic euthrerians (placentals, in black). Later taxa in light gray. All taxa more primitive than Mesozoic taxa were likely also present in the Jurassic and Cretaceous. None appear after Onychodectes. Madagascar separated from Africa 165-135 mya, deep into the Cretaceous with a population of tenrecs attached. No rafting was necessary.
The above represents what a robust cladogram is capable of, helping workers determine the likelihood of certain clades appearing in certain strata, before their discovery therein, based on their genesis, not their widest radiation or eventual reduction and extinction. In other words, we might expect sisters to basal primates, like adapids and lemurs, to be present in the Mesozoic, but not sisters to apes and hominids. We should expect sisters to all tree shrews and rodents to be recovered in Mesozoic strata. We should expect to see sisters to Caluromys, Vulpavus and other small arboreal therians/carnivorans in Mesozoic strata, but not cat, dog and bear sisters.
References Smith T 2011. Contribution of Asia to the evolution and paleobiogeography of the earliest modern mammals. Bulletin des séances- Académie royale des sciences d’outre-mer. Meded. Zitt. K. Acad. Overzeese Wet.57: 293-305
If you’re interested in dueling cladograms today you’ll see how two other cladograms (Figs. 1, 2) compete with the large reptile tree (LRT, subset Fig. 3, 1337 taxa) focusing on Metatheria/Marsupialia.
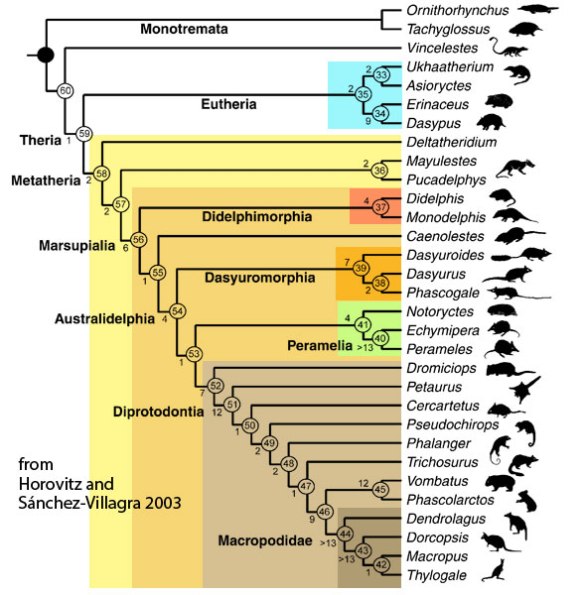
Figure 1. Marsupial cladogram based on Horovitz and xx 2006. Very few fossil taxa are present here.
Horovitz and Sánchez-Villagra 2003 (Fig. 1) used a trait analysis of mostly extant taxa to create their cladogram. Many prototherian and metatherian fossil taxa are missing here compared to the LRT (Fig. 3) where wombats and kangaroos have separate origins (Dasyurus is a last common ancestor), contra Horovitz and Sánchez-Villagra. And where is Sarcophilus (the Tasmanian devil)?
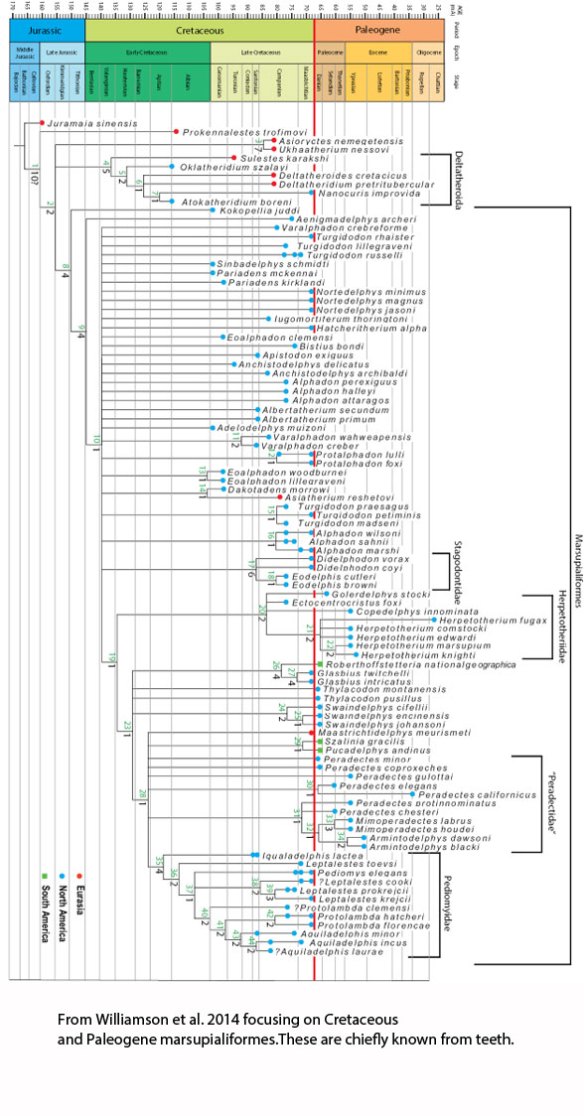
Figure 2. Marsupialiformes by Williamson et al. 2014 focusing on Cretaceous and Paleogene forms. Most of these are teeth and jaws only. By contrast the LRT includes many extant and recently extinct taxa known from skeletons.
Williamson et al. 2014 (Fig. 2) also lack a large number of fossil taxa and appear to avoid extant taxa compared to the LRT (Fig. 3). And where is Ernanodon (the Palaeocene Tasmanian devil)?
Figure 3. Subset of the LRT focusing on basal Mammalia. Both fossil and extant taxa are included here.
Reconstructions
(Fig. 4) help confirm that a list of sister taxa in a cladogram really do look like they are related to one another and demonstrate a gradual accumulation of traits (in lieu of laboriously examining all the data scores and confirming their validity with a worldwide trip to visit all the fossils firsthand).
Figure 4. From yesterday’s post, a selection of Therian skulls leading to placentals. This is the ‘gradual accumulation of traits’ recovered by the LRT.
A few words of wisdom from Neil deGrasse Tyson “Cognitive bias is all the ways we fool ourselves. Science. The only point of the scientific method is to make sure you are not fooled into thinking that something is true that is not, or thinking that something is not true that is… that is the only point.”
To that end: Adding taxa to the LRT tests and strains the data within that creates the cladograms presented here (Fig. 3). Errors are exposed and corrections are made. Today, the LRT cladogram remains highly resolved. That pretty much invalidates past criticisms. Today’s criticisms of those who think they are following the scientific method (Figs. 1, 2) are meant to get these authors to add relevant taxa, consider other candidates and options, re-examine their recovered trees, so that when they think something is true it will pass all tests and achieve confirmation and consensus that it is true… until the next study comes along to challenge it.
References Horovitz I and Sánchez-Villagra MR 2003. A morphological analysis of marsupial mammal higher-level phylogenetic relationships. Cladistics 19: 181–112. doi: 10.1111/j.1096-0031.2003.tb00363.x PDF Williamson TE and Brusatte SL and Wilson GP 2014. The origin and early evolution of metatherian mammals: the Cretaceous record. Zookeys. 2014; (465): 1–76.
Published online 2014 Dec 17. doi: [10.3897/zookeys.465.8178]
PMCID: PMC4284630 PMID: 25589872
There has been a traditional disconnect in mammalian paleontology regarding the two transitions between the egg-laying Prototheria, the pouched Metatheria, and the pouch-less Eutheria. So far as I can tell, only the large reptile tree (LRT, 1334 taxa; Fig. 2) has documented how and which species form transitional links in this chain of mammal evolution (Fig. 1). At present, and for the foreseeable future, didelphids, like Didelphis (the Virginia opossum), Monodelphis (the gray short-tailed opossum) and Caluromys (the wooly opossum) occupy basal nodes at large radiations of metatherians and eutherians in the LRT…hence the title of this post.
Figure 1. A selection of basal mammal skulls leading to placentals. This is the ‘gradual accumulation of traits’ recovered by the LRT. A third of these are traditional didelphids. Or two-thirds of these are cladistic didelphids. And, if so, then we humans are also didelphids. Haplodectes (IVPP V5235) nests as the basal primate in the LRT.
Traditionally Didelphidae has been a clade restricted to the opossums without any insight to their eventual descendants… the rest of the marsupials and us placentals. That’s why several mousy and not-so-mousy ‘possums have been added to the LRT recently, to more precisely recover evolutionary patterns in deep time. Amazing that our more or less direct ancestors are still with us today, sometimes hidden in Amazon forests, other times raiding our backyard trashcans and tentatively crossing our highways and byways.
Figure 1. Subset of the LRT focusing on basal Mammalia after the addition of several marsupials. Red taxa are represented by only a few bones, like mandibles with teeth. Note the proximity of traditional creodonts to the basal placental clade, Carnivora, basal members of which are small, arboreal and opossum-like.
A new taxon in the LRT is Thylophorops considered by Goin et al. 2009 to be the largest didelphid. Unfortunately, in the LRT, Thylophorops does not nest with Didelphis, but with Oxyaena and Thylacinus (Fig. 2)… themselves descendants of Didelphis with cat-like and wolf-like traits respectively.
Wikipedia reports, “Thylophorops species (as well as several other contemporary opossum genera) show a high degree of speciation towards carnivory compared to the still living didelphines. Their premolar and molar teeth were proportionally larger than those of living opossums and their grinding facets imply a more dedicated shearing action; these have been interpreted as “omnivory leading towards carnivory” in Goin et al. 2009.”
Figure 3. Crowned as the largest didelphid (by not much actually, but it is a juvenile) Thylophorops lorenzini nests between leopard-like Oxyaena and wolf-like Thylacinus in the LRT. All are shown to scale here.
Unfortunately
there is no reference in Goin et al. to either Oxyaena or Thylacinus. So… taxon exclusion is still an issue with the Goin et al. taxon list. Such problems are largely resolved in the LRT, which tests all possible candidates, and even dozens of fringe candidates that no one else considers, recovering a fully resolved tree based on traits and taxa that extend back to Devonian tetrapods, ultimately relating all descendants to one another.
References Ameghino F 1908. Las formaciones sedimentarias de la región litoral de Mar del Plata y Chapadmalal part 2 Goin FJ, Zimicz N, de los Reyes M, Soibelzon L 2009. A new large didelphid of the genus Thylophorops (Mammalia: Didelphimorphia: Didelphidae), from the late Tertiary of the Pampean Region (Argentina). Zootaxa. 2005: 35–46.
Maybe best known
from old Warner Brothers cartoons, (Fig. 1), the Tasmanian devil (Sarcophilus harrisii; Boitard 1841; up to 65 cm long) has long been considered the largest living and most carnivorous member of the Dasyuridae (typically insectivorous mouse-to-dog-sized marsupials, but see below).
Figure 1. The Warner Brothers version of the Tasmanian devil, affectionately called ‘Taz’ is known to eat a long list of animals…especially rabbits!
Typically
a Tasmanian devil will consume about 15% of body weight in food each day. It also makes ‘unearthly screams, coughs and growls’. The dog-like animal has red ears, wide jaws and big sharp teeth that it likes to display. It can sit on its haunches, like a raccoon. Seems to have been the wolverine from down under. No wonder Warner Brothers made such a gluttonous cartoon character! Here’s a 3:12 minute NatGeo special on the Tasmanian devil.
Sarcophilus laniarius ((originally Didelphis ursina Harris 1807) Owen 1839) is the extant Tasmanian devil, traditionally considered the largest dasyurid. Following the extinction of the taller, wolf-like Thylacinus in 1936 Sarcophilus became the largest living carnivorous marsupial.
Figure 2. Tasmanian devil (Sarcophilus) skeleton.
According to the Australian Parks and Wildlife webpage (below) “The famous gape or yawn of the Tasmanian devil that looks so threatening, can be misleading. This display is performed more from fear and uncertainty than from aggression.”
Figure 3. Sacrophilus skull from Digimorph.org and used with permission. Colors added.
Goodbye Marsupialiformes! In their study of Didelphodon, Wilson et al. 2016 employed the clade Marsupialiformes (Vullo and Gheerbrant 2009) for taxa not included within Marsupialia (living metatheres) and Metatheria. One of the problems with the Wilson et al. taxon list was… taxon exclusion. No placentals were included. The large reptile tree (LRT, 1134 taxa) documents the need for adding placentals and fossil taxa because placentals diverged from arboreal carnivorous metatheres, like Caluromys in the LRT, as we learned earlier here. The opossum, Didelphis, nests as a derived marsupial in Wilson et al. It nests as a basal metathere in the LRT.
Here’s another Tasmanian Devil YouTube video:
Link to further details and sound file from the Australian Parks and Wildlife webpage.
Over the holidays I have been busily adding taxa to the LRT, mostly small, mouse-like opossums, in order to iron out the phylogenetic issues that arose there. The corrections are solving problems that we will review over the next few days. Today, I can tell you that the Tasmanian devil (genus: Sarcophilus) nests between the smaller extant quoll (genus: Dasyurus, no surprise there) and the devil-sized, formerly enigmatic Palaeocene Ernanodon (Fig. 4). If I am not mistaken, Sarcophilus and Ernanodon have never been tested together until now. This pushes the origin of pugnacious little Sarcophilus back to the K-T boundary, and possibly much deeper in time and not restricted to Tasmania or Australia where Sarcophilus fossils have been found that are less than 100,000 years old.
Molecular evidence
suggests devils split from quolls between 10 and 15 million years ago (Krajewski and Westerman 2003).
Figure 4. Now nesting more precisely and closely with Sarcophilus is Ernanodon from the Palaeocene of China. This enigma taxon was traditionally linked to anteaters, then pangolins. Perhaps Sarcophilus and Ernanodon have never been tested together until now. Note the difference in the feet between the museum mount and the drawing inserts. The inserts look more like those of sister taxa. Repairing such problems is what has to be done to create a high resolution cladogram.
References Boitard P 1842. L’Ursin de Harris. Le Jardin des plantes: Description et mœurs des mammifères de la Ménagerie et du Muséum d’histoire naturelle. Paris: Gustave Barba. p. 204. Harris GP 1807. Description of two new Species of Didelphis from Van Diemen’s Land. Transactions of the Linnean Society of London. 9: 174–78. Krajewski C and Westerman M 2003. Molecular Systematics of Dasyuromorpha. In Jones, Menna; Dickman, Chris; Archer, Mike. Predators with Pouches: The Biology of Carnivorous Marsupials. Collingwood, Victoria: CSIRO Publishing. p. 16. Vullo R and Gheerbrant E 2009. The oldest modern therian mammal from Europe and its bearing on stem marsupial paleobiogeography. Proceedings of the National Academy of Sciences106(47):19910-19915. Wilson GP, Eddale EG, Hoganson JW, Calede JJ and Vander Linden A 2016. A large carnivorous mammal from the Late Cretaceous and the North American origin of marsupials. Nature Communications 7:13734 PDF
According to Wikipedia “Marsupialiformes [ Marsupialia sensu Kielan- Jaworoswka et al.] is erected to account for the crown group Marsupialia (extant marsupials and related extinct fossil taxa) plus all stem marsupialiform taxa that are more closely related to them, as their sister taxa, than to Deltatheroida and basal Metatheria. Basal marsupialiforms, such as the North American taxa from the Early/Late Cretaceous, include the stem groups of the crown marsupials. Basal marsupialiforms include primitive Cretaceous taxa previously gathered in the paraphyletic taxon ‘‘Ameridelphia’’.
Metatheria includes Marsupialiformes and Deltatheroida (presumed sister groups), plus basal metatherians, such as Sinodelphys. Unfortunately Sinodelphys nests with the basal prototherian Megazostrodon in the LRT, so that raises a red flag.