Updated February 15, 2014 with the addition of several basal snakes to the large reptile tree (498 taxa) that unites terrestrial snakes and burrowing snakes in one clade derived from basal gekkotans.
Caldwell et al. (2015)
add 70 million years to the oldest known snakes previously known. Now snake origins go back to the Middle Jurassic/Early Cretaceous. These new fossils are not complete fossils — and they might have had legs. The few parts that are known (Fig. 1) indeed look like snake parts.
This is the paper for which Caldwell and Apesteguía asked me to delete and postpone my work on Jucarasepsp). However, as near as I can tell after reading the paper and supplementary information, Jucaraseps is not mentioned.

Figure 1. Maxillae from several Mid-to-Late Jurassic snakes discovered by Caldwell et al. 2015, including Parviraptor, Diabolosphis and Portualophis.. These are compared to Lanthanotus, an extant limbed precursor to burrowing snakes, like Cylindrophis. Also shown are Eichstaettisaurus, a limbed varanid precursor to terrestrial snakes along with the skulls of Adriosaurus, an elongate limbed pre-snake and Pachyrhachis, a Late Cretaceous snake with vestigial hind limbs and no forelimbs or pectoral girdle.
Unfortunately,
in the Caldwell et al. paper their phylogenetic analysis nests snakes with legless amphisbaenids (including Dibamus at their base), odd Sineoamphisbaena, mosasaurs (ncluding Adriosaurus at their base) and varanids (including Heloderma and Lanthanotus) as successively more distant outgroups. It may be that the legless traits swamped any skull differences — and they are legion — yet with many, many convergences.
In the large reptile tree Sineoamphisbaena and a variety of proto-amphisbaenids are derived from skinkomorphs, distinct from varanids, helodermatids and their allies. And certainly distinct from snakes.
The Caldwell et al. paper nested the odd to very odd burrowing snakes, such as tiny Leptotyphlops, in the midst of more traditional above ground snakes. In the large reptile tree Leptotyphlops nests as a very derived taxon.
Snakes are indeed difficult taxa to lump and split. There is a long history of attempts at nesting snakes within the Lepidosauria, many with distinctly different trees and distinct snake origin hypotheses.
An alternate tree
The large reptile tree did not recover the same topology. And the nesting of snakes has taken an odd twist from what I presented over the last four years.
Snakes evolved from the marine taxon, Adriosaurus, which evolved from the terrestrial taxa Jucaraseps and Eichstaettisaurus in order of increasing distance. That hasn’t changed.
What has changed is snake ancestors no longer evolve from basal varanids (which also gave rise to mosasaurs). So Adriosaurus is not a varanid in the new large reptile tree.
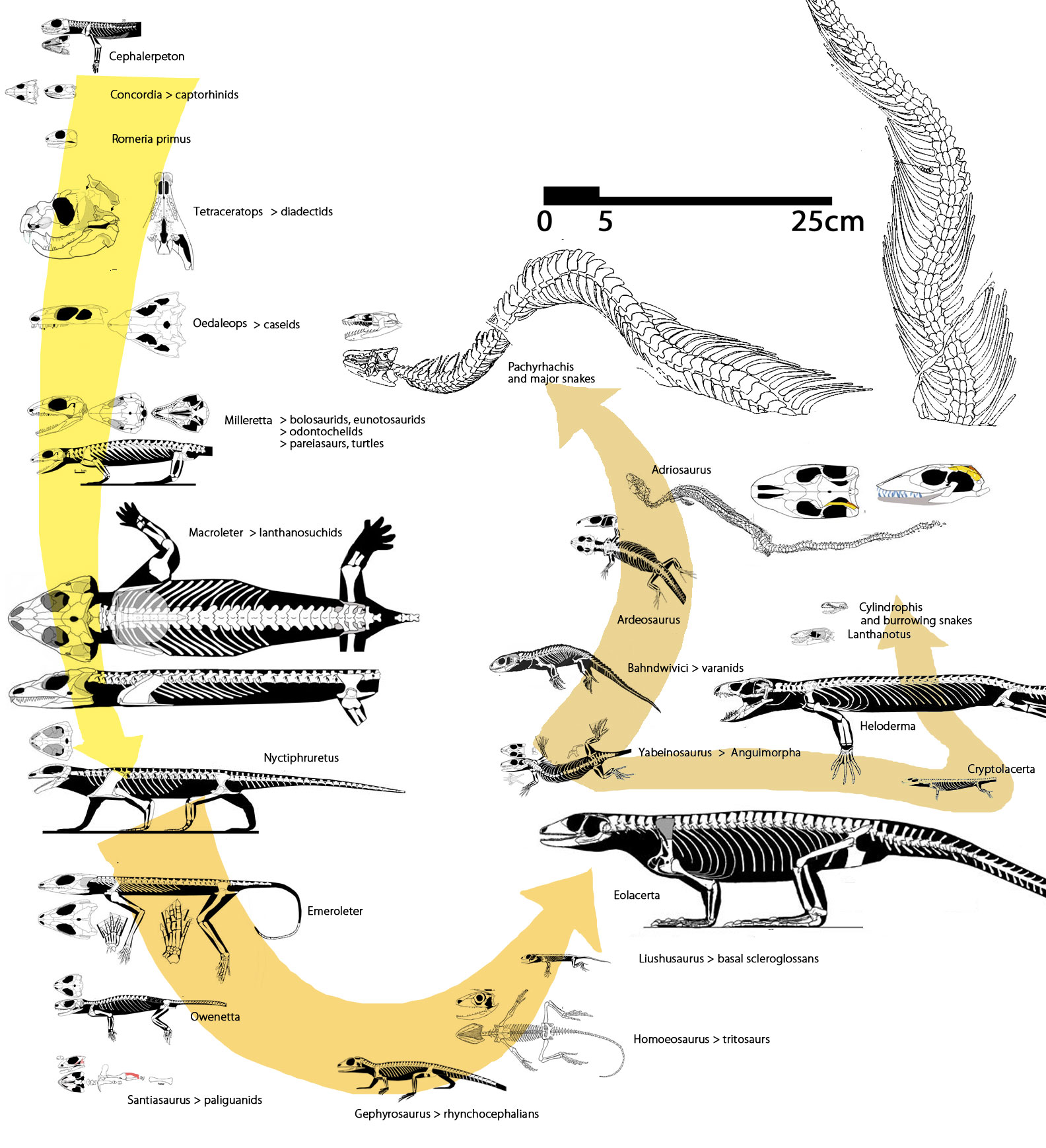
Now, according to the large reptile tree, the last common ancestor of traditional snakes through Eichstaettisaurus and Ardeosaurus is Tchingisaurus. So snakes are indeed an ancient clade, likely going back to the Triassic and the lineage of their ancestry goes back to the Middle Permian. Remember, the Ascendonanus nests within the iguanids, and it lived in the Early Permian, the earliest squamate specimen in this taxon list (Fig. 2), and it was more derived than the last common ancestor of skinks, geckos, varanids, snakes and helodermatids. This nesting also finds support in earlier nestings by Conrad (2008) that nested the snake ancestors Ardeosaurus and Eichstaettisaurus tentatively with geckos. In the large reptile tree these taxa now nest closer to geckos than to varanids.

Figure 2. Subset of the Large reptile tree focusing on snake origins within the Squamata.
Ardeosaurus and Eichstaettisaurus were Late Jurassic with origins probably much earlier. Jucaraseps was Early Cretaceous.
The longevity of several lepidosaur clades, including the extant Sphenodon, is remarkably distinct from the relatively shorter appearances of most archosauromorph, archosauriform, protorosaur and enaliosaur clades.
More on this topic later. I’ll rearrange the pages at reptileevolution.com to reflect these changes and add the taxa that made that happen. Science is a series of improvements.
References
Caldwell MW, Nyam RL, Placi A and Apesteguía S 2015. The oldest known snakes from the Middle Jurassic-Lower Cretaceous provide insights on snake evolution. Nature Communications 6: 5996.[6996]
Conrad JL 2008. Phylogeny and systematics of Squamata (Reptilia) based on morphology. Bulletin of the American Museum of Natural History 310. online here.